


INTRODUCTION
Atopic dermatitis (AD) is a general allergic disease that is characterized by eczematous and pruritic skin lesions secondary to skin barrier dysfunction. It occurs during infancy and childhood [1,2]. However, it may also occur in the middle age and persist for life. AD affects approxi-mately 14-20% of children and 1-4% of adults, and its prevalence contin-ues to increase, particularly in low-income countries [3]. AD is a com-plex disease, involving several factors such as an abnormal immune system, genetic disorders, and environmental factors [4-7]. Recent research has provided greater insights into the molecular and immunological mechanisms involved in AD [2].
AD can be caused by imbalances in regulatory T cells; patients with AD exhibit increased differentiation of helper T (Th)-2 cytokines, such as interleukin (IL)-4 and IL-5, which inhibit Th1 cell production [8]. Th2 cell-mediated and Th1 cell-mediated lesions have been reported in the acute and chronic phases of AD, respectively [9,10]. Furthermore, tumor necrosis factor-alpha (TNF-α) and interferon-gamma (IFN-γ) have also been reported to play important roles in acute and chronic AD [11,12]. Thymic stromal lymphopoietin (TSLP) is an epithelial-derived cytokine; its abnormal expression has been reported to be related to atopic diseases [13-15].
Generally, moisturizers and topical steroids that maintain membrane moisture levels and suppress the inflammatory response are prescribed for the treatment of AD. However, they also have some side effects [16]. Therefore, several investigators have focused on developing natural anti-inflammatory agents with fewer side effects [17]. Copper peptides (CPs) are naturally occurring complexes that have been used for a variety of skin-related purposes [18,19]. They have been reported to have a significant effect on collagen production [18]. CP-containing cosmetics im-prove the condition of the skin and decrease visible signs of aging by in-creasing the epidermal thickness, skin elasticity, and skin density [19]. However, very little is known about the effects of CP on AD. In the current study, we determined the inhibitory effect of CP on AD induced by house dust mite extract (HDM) and 2,4-dinitrochlorobenzene (DNCB) in BALB/c mice. For this purpose, we examined the dermal and epidermal ear thickness as well as the serum immunoglobulin E (IgE) levels, mast cell infiltration levels, and mRNA gene expression levels of pathogenic cytokines in the ear tissue. The expression of IL-4 mRNA in CD4+ T lymphocytes in the draining lymph nodes (dLNs), as well as the weight and size of these dLNs, were also determined. We further investigated the immunomodulatory and anti-inflammatory effects and the action-related mechanisms of CP in vivo and in vitro. We confirmed the inhibitory effects of CP on AD symptoms by using the AD-skin like mouse model BALB/c in the in vivo study, and clarified the effects exerted by CP on certain signaling pathways within the human keratinocyte (HaCaT) cell line in the in vitro study.
METHODS
1. Materials
TRIzol reagent for RNA extraction was bought from Invitrogen (Carlsbad, CA, USA). Primary antibodies and peroxidase-conjugated secondary antibodies were bought from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). All other reagents were of the highest grade commercially available at the time of the study.
2. Procedure for CP
CP was synthesized chemically. Briefly, CP was formulated in buffer (Bioprogen, Daejeon, Republic of Korea) and confirmed by the appearance of a sky blue solution (pH 6.8-7.0).
3. Animals
Seven-week-old female BALB/c mice were bought from Samtako (Osan, Republic of Korea) and housed under specific pathogen-free con-ditions. All experiments were approved by the Institutional Animal Care and Use Committee of Konkuk University (approval number: KU14012).
4. Induction of AD lesions in the ear
AD was induced in the mice by repeated local exposure of their ears to HDM extract and DNCB, as described previously [20]. The mice were separated into the following five groups: control, AD-only, AD+CP10, AD+CP100, and AD+ceramide (CERA; positive control). The surfaces of both earlobes were stripped five times with surgical tape (Nichiban, Tokyo, Japan). Thereafter, 20 µL of 1% DNCB was applied to each ear, followed by 20 µL of HDM (10 mg/mL) applied similarly 4 days later. HDM or DNCB treatment was administered once per week for 4 weeks. The mice received CP, CERA, or both throughout the 4-week AD induction regimen. Ear thickness was measured 24 hours after HDM or DNCB application using a dial thickness gauge (Kori Seiki MFG, Co., Japan). At days 14 and 28, blood samples were collected via an orbital puncture and centrifuged to yield plasma samples (*), which were stored at −70°C for further analyses. After blood collection, the ears were re-moved and subjected to a histopathological analysis. The IgE levels were measured at 28 days after the first induction using an IgE enzyme-linked immunoassay kit (Bethyl Laboratories, Inc., Montgomery, TX, USA) in accordance with the manufacturer's instructions.
5. Histological observations
The ears were fixed in 10% paraformaldehyde for 16 hours and embedded in paraffin. Thin sections of 5 µm were stained with hematoxylin and eosin. Dermal and epidermal thickness were measured using microscopy. To evaluate mast cell infiltration, the skin sections were stained with 0.01% toluidine blue, and the number of mast cells was counted in five randomly chosen fields of view.
6. Real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (PCR) was performed using Thermal Cycler Dice TP850 (Takarabio Inc., Shiga, Japan) in accordance with the manufacturer's protocol. For each group, the total RNA was isolated from the ear tissue or the CD4+ T lymphocytes in the dLNs. To determine IL-2 expression, the cells were stimulated with phorbol my-ristate acetate+A23187 for 24 hours. The resulting supernatant was collected and analyzed quantitively for the IL-2 concentration. The PCR conditions were similar to those described previously [20]. Briefly, 2 μL of cDNA (100 ng), 1 μL of the sense and antisense primer solution (0.4 µM), 12.5 μL of SYBR Premix Ex Taq (Takarabio Inc.), and 9.5 μL of dH2O were mixed to obtain a 25 μL reaction mixture in each reaction tube. The following mouse primer pairs were used for PCR: mouse TNF-α, 5′-AAG CCT GTA GCC CAC GTC GTA-3′ and 5′-GGC ACC ACT AGT TGG TTG TCT TTG-3′; mouse IFN-γ, 5′-TCA AGT GGC ATA GAT GTG GAA GAA-3′ and 5′-TGG CTC TGC AGG ATT TTC ATG-3′; mouse IL-4, 5′-ACA GGA GAA GGG ACG CCA T-3′ and 5′-GAA GCC GTA CAG ACG AGC TCA-3′; mouse IL-17, 5′-CGG AGG CCA ACA ACA AGA GA-3′ and 5′-CTC CCT CAC TGT GGC TCT TC-3′; mouse IL-31, 5′-CCT ACC AGA CCA AGG TCA AC-3′ and 5′-AGG GGG TAA TAA AGG GAT TG-3′; mouse TSLP, 5′-TCC TCT GAA GAC CTG ACC-3′ and 5′-TCT CCT TTC TCC CTA ATC CTC-3′; and mouse GAPDH, 5′-GCA CAG TCA AGG CCG AGA AT-3′ and 5′-GCC TTC TCC ATG GTG GTG AA-3′. The following human primer pairs were used for PCR: human TNF-α, 5′-CCT ACC AGA CCA ACC TCA AC-3′ and 5′-AGG GGG TAA TAA AGG GGA TTG-3′; human IL-6, 5′-AAA AGA GCA CTG CCA AGG AA-3′ and 5′-ATC TGA GGT GCC CAT GCT AC-3′; human IL-1ϐ, 5′-GGA TAT GGA GCA ACA AGT GG-3′ and 5′-ATG TAC CAG TTG GGG AAC TG-3′; human TSLP, 5′-TAG CAA TCG GCC ACA TTG CCT-3′ and 5′-GAA GCG ACG CCA CAA TCC TTG-3′; and human GAPDH, 5′-CGG AGT CAA AGG ATT TGG TCG TAT-3′ and 5′-AGC CTT CTC CAT GGT GGT GAA CAC-3′. The amplification conditions were as follows: 10 seconds at 95°C, 40 cycles of 5 seconds at 95°C and 30 seconds at 60°C, 15 seconds at 95°C, 30 seconds at 60°C, and 15 seconds at 95°C. The mRNA levels of the target genes relative to GAPDH were normalized using the following formula (Ct: threshold PCR cycle value): relative mRNA gene expression=2–(Ct of target gene−Ct of GAPDH). In each sample, the mRNA expression level of the analyzed gene was normalized to that of GAPDH and presented as the relative mRNA level.
7. Cell culture
The HaCaT cells were cultured in Dulbecco's modified Eagle's medium (supplemented with 10% fetal bovine serum as well as penicillin/strepto-mycin [10 µg/100 mL]) in 5% CO2 at 37°C. For the migration assay, the HaCaT cells were pre-incubated with TNF-α (10 ng/mL, PeptroTech, Rocky Hill, NJ, USA) and IFN-γ (10 ng/mL, PeptroTech, Rocky Hill, NJ, USA) for stimulation. The cells were then harvested after 6 hours, and the protein and total RNA were isolated. The morphology of the HaCaT cells was examined using a phase-contrast microscope (Olympus CX41; Olympus, Tokyo, Japan).
8. Statistical analysis
Experiments were performed independently at least three or four times on different days. Statistical analysis was performed using the SAS statis-tical software (SAS Institute, Cary, NC, USA). Multiple group data were analyzed using a one-way analysis of variance followed by the Dunnett's multiple range test. All results are expressed as mean±standard deviation of comparative fold differences. Data are representative of three indepen-dent experiments. The threshold for significance was set at p<.05.
RESULTS
1. Chemical characterization of CP and induced AD in the BALB/c mouse ear
Fig. 1 is a schematic depiction of the induction and CP-based treatment of AD in the BABLB/c mouse model. AD was induced in the mice ears by alternately applying HDM extract and 1% DNCB on both earlobes for 5 weeks; the effects of CP treatment were evaluated. A week prior to exposure, induction with HDM extract and DNCB were tested in a preliminary experiment (Fig. 1A). The absorbance curves of copper, peptide, and copper peptide are provided in Fig. 1. Presence of CP was confirmed by the appearance of a sky blue solution (pH 6.8-7.0), formulated with buffer (Fig. 1B).
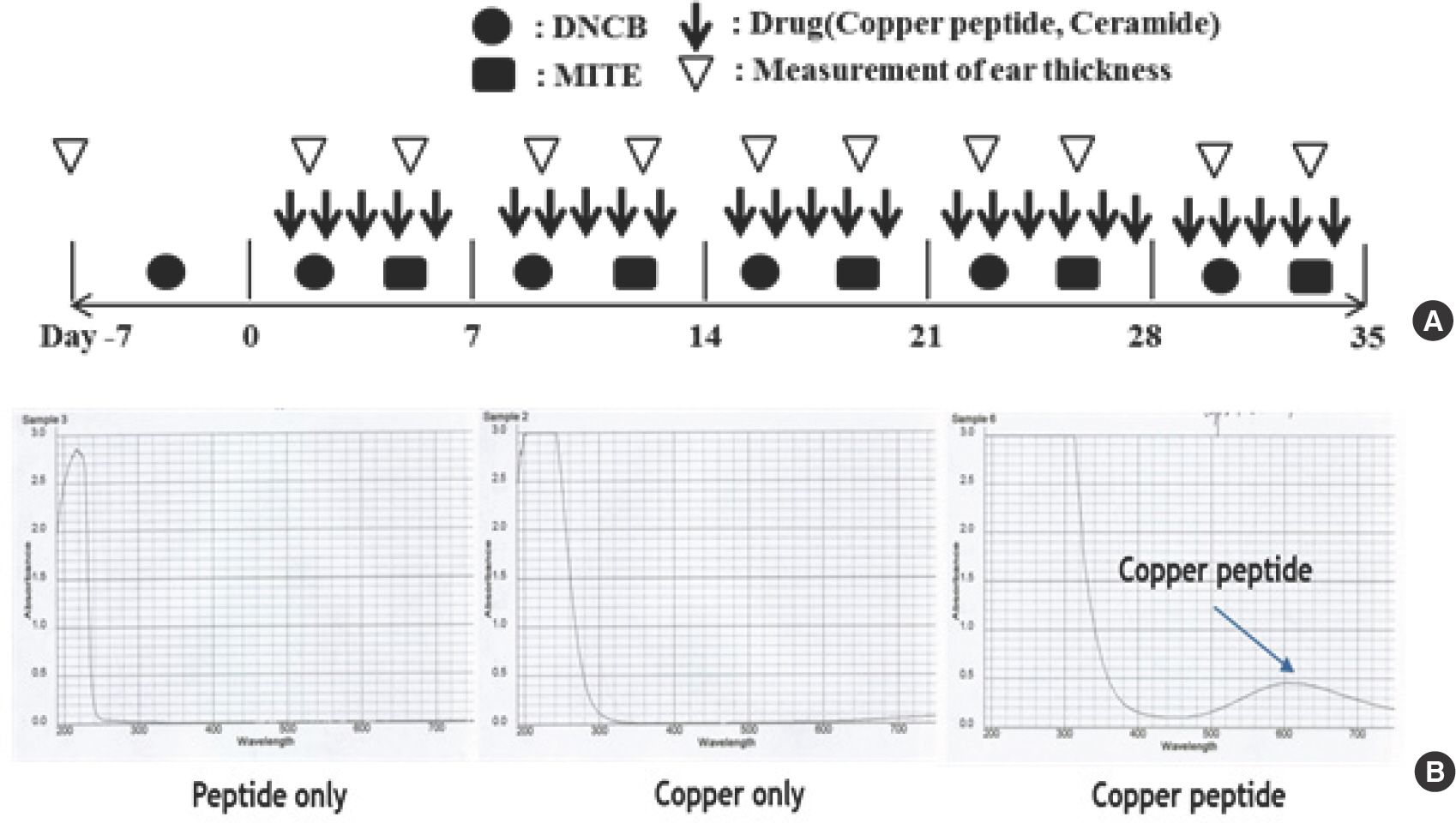
2. CP reduces BALB/c mouse ear thickness and serum IgE level
HDM/DNCB application increased the ear tissue thickness and AD severity in BALB/c mice. The ear thickness and the extent of mast cell and T cell infiltration in the ear tissue were examined after topical CP treatment. Compared to mice untreated with CP (AD-only group), mice with AD who were treated with CP exhibited a reduced ear tissue thickness and AD severity (*based on mast cell and T cell infiltration; Fig. 2A). The number of lesions on the mouse ears at __ days after the start of AD induction were lower in mice from all CP-treated groups than in mice from the AD-only group (Fig. 2B).
Fig. 2.
Fig. 2.(A) Ear thickness was measured with a dial thickness gauge every 3 days after HDM extract or DNCB application. (B) Typical photographs of the mouse ears from each group obtained at 28 days after starting AD induction. CP, copper peptide; CON, control; CERA, ceramide; AD, atopic dermatitis; HDM, house dust mite; DCNB, 2,4-dinitrochlorobenzene.
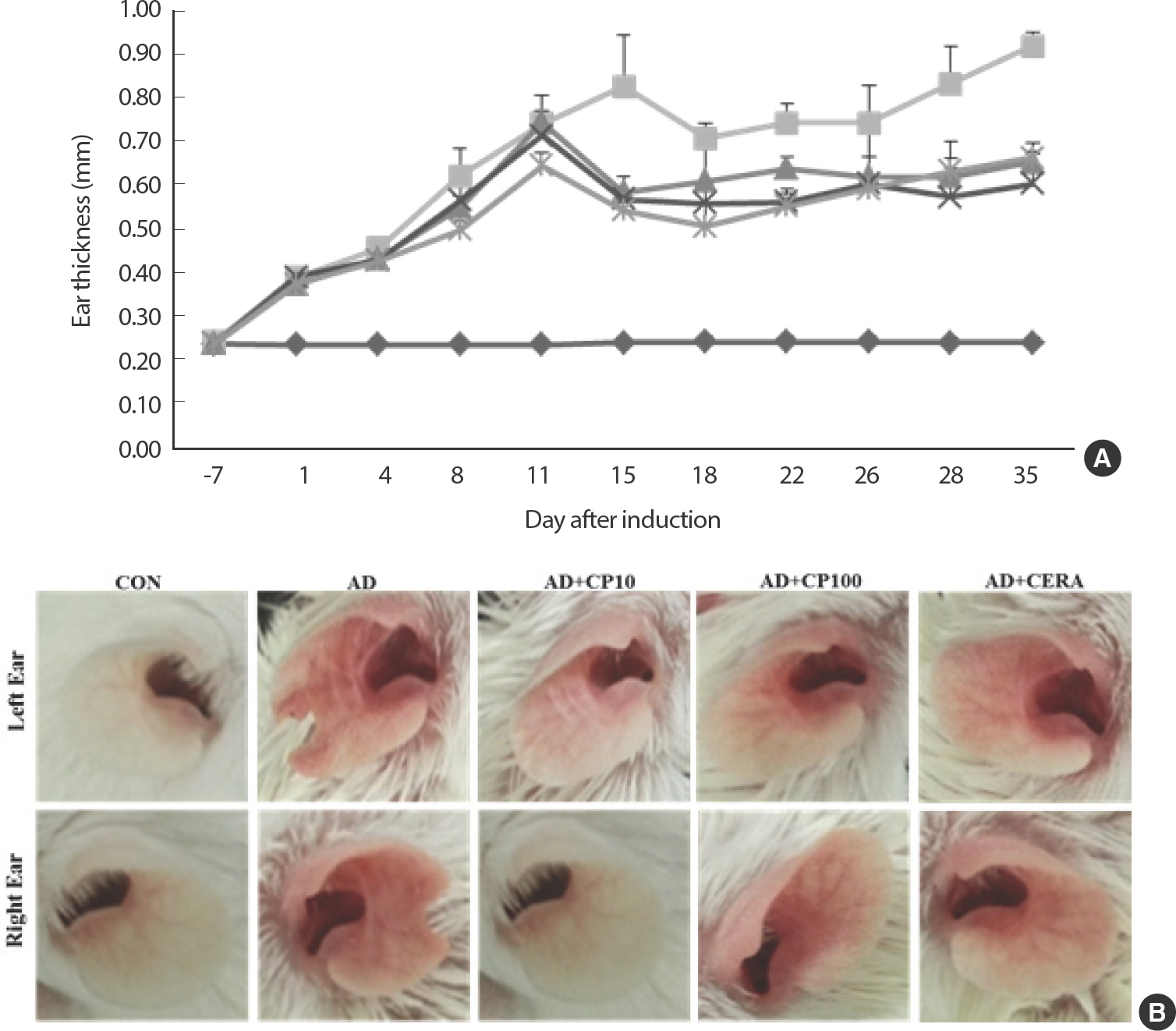
3. CP reduces immune cell infiltration and mouse ear tissue inflammation
Compared with the AD-only and AD-CERA groups, the CP-treated groups exhibited a significantly decreased number of infiltrated mast cells in a dose-depended manner (Fig. 3C, D). Furthermore, the effect of CP was more potent than that of CERA (i.e., the positive control). CP treatment also suppressed epidermal and dermal thickness (Fig. 3A, B). The serum IgE level was higher in mice with AD than in the control mice. However, the total serum IgE level was significantly lower in the CP-treat-ed mice than in the AD group mice in a concentration-dependent man-ner (Fig. 3E).
Fig. 3.
Fig. 3.Histopathological and serum analyses of the effect of CP on mast cell infiltration. Dermal and epidermal thickness were measured using microphotographs of eosin-and-hematoxylin-stained-tissue (A, B). Typical photomicrographs of ear sections stained with eosin or toluidine blue and hematoxylin (D). (C) The number of infiltrated mast cells was determined on the basis of toluidine blue staining. Blood samples were collected via orbital puncture on day 28. The plasma IgE levels were quantified by enzyme-linked immunosorbent assay. Data are presented as mean±SD from triplicate measurements. The original magnification was 100×CON. IgE, immunoglobulin E; CP, copper peptide; CON, control; CERA, ceramide; AD, atopic dermatitis; SD, standard deviation.* Indicates significant difference from the values of the HDM/DNCB-treated mice at p<.05. AD induced by HDM and DNCB treatment. The pictures shown are representative of each group (n=3–6).

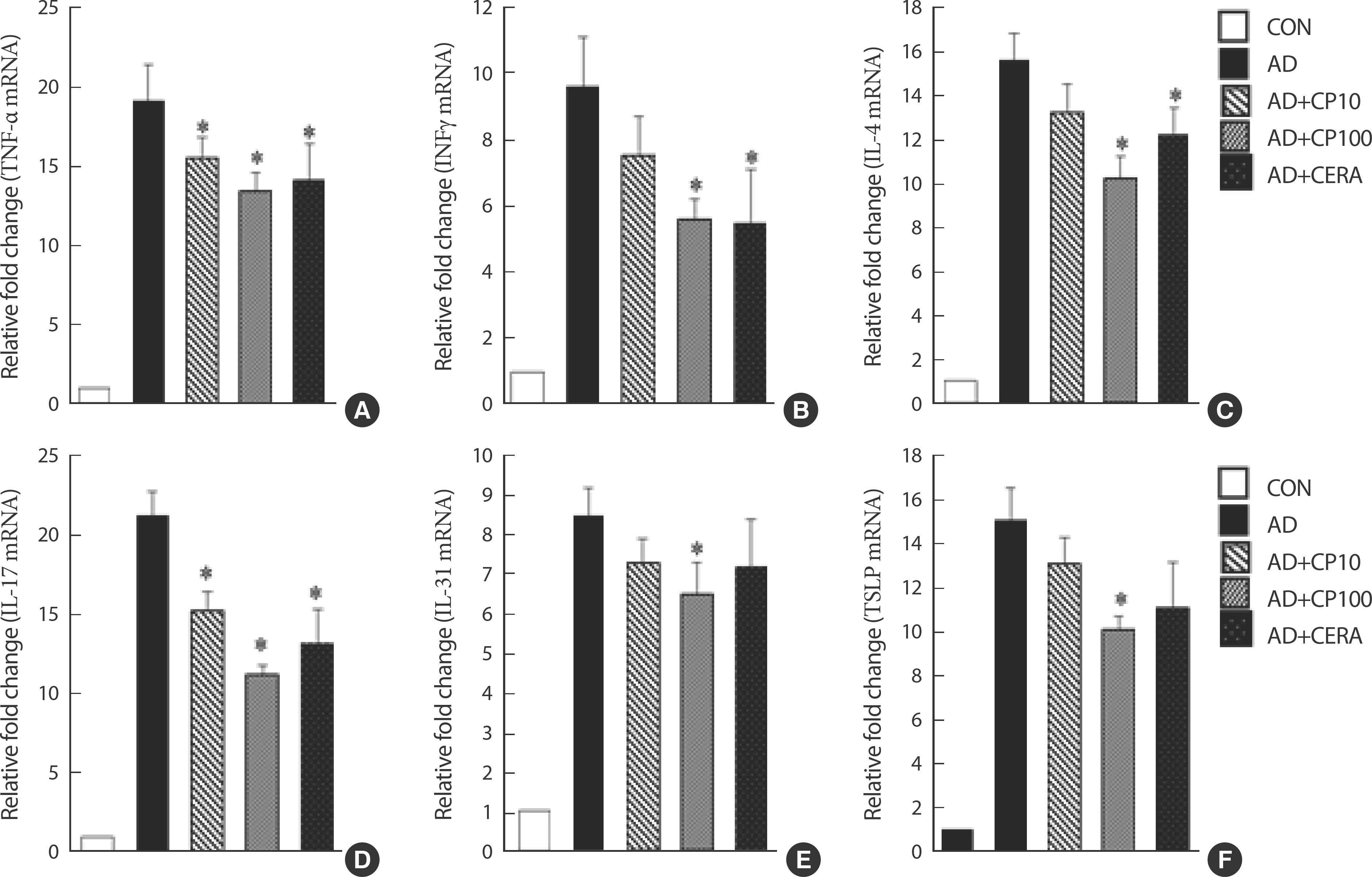
4. CP treatment downregulates mRNA gene expression of cytokines in the ear tissue
CP treatment significantly inhibited both Th1 and Th2 cytokines (Fig. 4). CP treatment decreased the expression of Th1-related cytokines (TNF-α and IFN-γ; Fig. 4A, B) and Th2-related cytokines (IL-4, IL-7, and IL-31) in the ear tissue (Fig. 4C-E). CP inhibited the expression of TSLP in a dose-dependent manner. Remarkably, the effects of CP100 were more robust than those of CERA (Fig. 4F).
Fig. 4.
Fig. 4.(A–F) Effect of CP on the mRNA expression of various pathogenic cytokines in the ear tissue of BALB/c mice. The ears were excised on day 28, and the total RNA was isolated. Quantitative real-time PCR was performed as described in the Methods. Data are presented as mean±SD of triplicate measurements. HDM, house dust mite; DNCB, 2,4-dinitrochlorobenzene; CP, copper peptide; CON, control; CERA, ceramide; AD, atopic dermatitis; SD, standard de-viation; PCR, polymerase chain reaction.* Indicates significant difference from the values of the HDM/DNCB-treated mice at p<.05. AD induced by HDM and DNCB treatment.
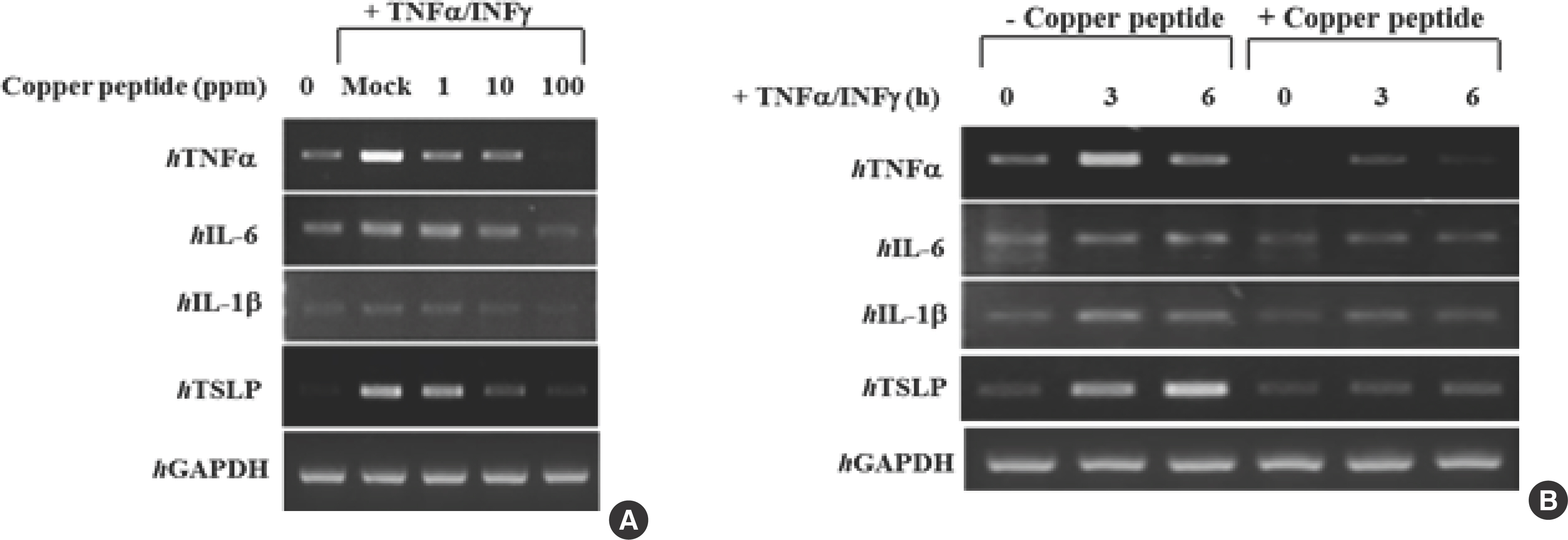
5. Inhibitory effect of CP on the inflammatory response in stimulated HaCaT keratinocytes
The ability of CP to block the effects of the proinflammatory cytokines TNF-α and IFNγ (10 ng/mL) in HaCaT keratinocytes was assessed using real-time PCR (Fig. 5). CP effectively decreased hTNF-α, hIL-1β, hIL-6, and hTSLP mRNA expression in IFN-γ/TNF-α-induced cells in a dose-dependent manner. This indicated that CP is capable of inhibiting proinflammatory cytokine production in human keratinocytes (Fig. 5A, B).
Fig. 5.
Fig. 5.HaCaT cells were stimulated by pre-incubation with TNF-α (10 ng/mL) and IFN-γ (10 ng/mL) (A). After 6 hours of stimulation, the cells were harvested, and the total RNA was isolated. Gene expression levels of proinflammatory cytokines (hTNFα, hIL-1β, hIL-6, and hTSLP) were measured using conventional real-time PCR (B). HaCaT, human keratinocyte cell line; TNF-α, tumor necrosis factor-alpha; IFN-γ, interferon-gamma; IL, interleukin; TSLP, thymic stromal lymphopoietin; PCR, polymerase chain reaction.
DISCUSSION
CP has a broad range of effects on aging and skin remodeling [21,22]. We evaluated the therapeutic effects of CP on AD in BALB/c mice (induced by alternately applying HDM and DNCB to both earlobes for 5 weeks). HDM/DNCB use significantly increased the number of AD lesions and the ear thickness. The mouse ear thickness and inflammatory cell infiltration, serum IgE levels, and mRNA expression of immunoreg-ulatory cytokines in the mouse ear tissue were examined after topical CP treatment. The number of lesions and ear thickness in the CP-treated mice was lower than that in the AD mice (Fig. 2A, B). Mast cells play an important role in the induction of inflammation. Activated mast cells release potent inflammatory mediators, such as cytokines and hista-mine. Thus, histological analysis of the atopic ears was performed to ex-plain the AD symptoms noted visually. The ears excised from each mice group were stained with toluidine blue or hematoxylin, and mast cell infiltration and dermal and epidermal thickening were examined under a microscope. Compared with the AD group, CP-treated groups showed a significantly decreased number of infiltrated mast cells in a dose-de-pended manner (Fig. 3C, D). Furthermore, the effect of CP was more potent than that of CERA. CP treatment also suppressed epidermal and dermal thickness (Fig. 3A, B). Elevated IgE levels are one of the important signs of AD progression [23]; a large body of studies have revealed that AD is a representative Th2 type immune disorder wherein the serum IgE levels and Th2 type cytokine expression are upregulated [24,25]. Next, IgE level was measured in each treatment group; as expected, the IgE level was higher in the AD mice than in the control mice. Compared to the AD group mice, the CP-treated mice exhibited significantly reduced total and HDM-specific IgE levels in a concentration-dependent manner (Fig. 3E). Interestingly, the therapeutic effects of CP were more potent than those of CERA.
Further, to determine whether CP exerted its effects mostly on the Th1 or Th2 response and to demonstrate the underlying mechanism of CP, we analyzed the mRNA gene expression levels of AD-related pathogenic cytokines in the ear tissues using real-time PCR. The mRNA expression of all the tested cytokines was upregulated in the ear tissue of AD mice. Topical CP treatment significantly inhibited both Th1 and Th2 cytokines (Fig. 4). The CP treatment decreased the expression of both Th1-related cytokines (TNF-α and IFN-γ; Fig. 4A, B) and Th2-related cytokines (IL-4, IL-7, and IL-31) in the ear tissue (Fig. 4C–E). TSLP is closely related to IL-7 and plays a considerable role in allergic diseases, including AD [26]. Therefore, we next investigated the expression of TSLP in the ear tissue. We found that CP inhibited TSLP expression in a dose-dependent man-ner. Remarkably, the effects of CP100 were more robust than those of CERA (Fig. 4F). Notably, Soumelis et al. [27] found high levels of TSLP in the skin of patients with AD, but not in unaffected skin. Furthermore, several researchers have investigated the biological effects of cytokines on keratinocytes: they have concluded that keratinocytes are markers for a specific set of cytokines involved in skin inflammation [28]. Besides, IL family members have also been distinctly identified as significant controllers of the epidermal functions; their receptors have also been noticed on keratinocytes [29]. In the present study, the effect of CP on HaCaT keratinocytes induced by proinflammatory mediators TNF-α (10 ng/mL) and IFN-γ (10 ng/mL) was assessed using RT-PCR (Fig. 5). CP effectively decreased hTNF-α, hIL-1β, hIL-6, and hTSLP mRNA expression in the IFN-γ/TNF-α-induced cells in a dose-dependent manner. This indicates that CP is capable of inhibiting proinflammatory cytokine production in human keratinocytes (Fig. 5A, B). Furthermore, Bogiatzi et al. [30] also reported that high levels of Th2 cytokines and proinflammatory sub-stances in the affected skin cooperate to induce TSLP production by keratinocytes; this implies the role of an inflammatory pathway. Our findings are also consistent with these observations.
CONCLUSIONS
Herein, we investigated the anti-AD potential of CP using a representative animal model of AD. Representative application of CP significantly decreased the severity of AD in terms of the histopathological symptoms, IgE production, and mRNA expression of pathogenic cytokines in HDM/DNCB-induced BALB/c mice and human keratinocytes. Taken together, our results suggest that CP is effective in inhibiting the AD-like phenotype by regulating the inflammatory response.


