Melatonin as an Antioxidant Supplement in Athletes: A Literature Review of Current Evidence
Article information

Abstract
PURPOSE
Melatonin (MT; N-acetyl-5-methoxytryptamine) is synthesized and released during the night in specialized pineal gland cells. Among its variety of physiological properties, recent research indicates that MT has both antioxidant and anti-inflammatory actions. The aim of this literature review was to summarize recent evidence that describes the effects of MT on the muscle function and preventive role of MT on exercise-induced muscle damage.
METHODS
This review included previous research using the PubMed, Science Direct, and Google Scholar databases.
RESULTS
We discussed the molecular structure and biological function of MT and the potential role of this hormone in antioxidant and anti-inflammatory processes. These activities have been studied in relation to the protection of muscle function against oxidative damage. In addition, MT is reported to have positive effects on muscle damage, lipid metabolism, and inflammatory responses in well-trained athletes following exercise training. Moreover, the potential beneficial effects of melatonin and mechanisms related to performance were revealed through improved sleep quality, muscle damage, and antioxidant levels in trained athletes.
CONCLUSIONS
Finally, this review suggested that possibilities of MT as a supplementation for athletes; however, further research is required to investigate the specific mechanisms involved, the dose and duration of use, and the beneficial and detrimental effects of MT on athletic performance.
INTRODUCTION
Vigorous exercise training and submaximal muscle contraction ele-vates the generation of reactive oxygen species (ROS) and reactive nitrogen species (RNS), inducing oxidative stress in skeletal muscle [1]. In-creased ROS-induced oxidative stress negatively affects cellular components, including proteins and DNA, which can result in muscle damage and fatigue [2]. On the other hand, recent evidence emphasizes the significance of redox signaling on reactions of skeletal muscle following the exercise training, including the increase of mitochondrial biogenesis and endogenous antioxidants enzymes as well as other health benefits such as glucose metabolism, growth factor signaling and immune system [3,4]. In this respect, continuous repetitive training enhances antioxidant capacity and reduces oxidative damage [5].
Growing evidence shows that elite athletes can be susceptible to oxidative stress during frequent high-intensity training sessions, which results in greater oxygen consumption and increased ROS production [6]. In addition, insufficient rest and lack of proper recovery between training sessions preclude adequate repair and adaptation after exercise-induced muscle damage, which can lead to cumulative oxidative stress. In-creased ROS production can induce the activation of cellular transcription factors such as NF-κB which control systemic inflammatory re-sponse [7]. Thus, maintaining a balanced ROS level is particularly im-portant for minimizing muscle damage for athletes [8].
Antioxidants play a role in protection against the harmful effects of ROS by exercise training. Recently, melatonin (MT; N-acetyl-5-methoxy-tryptamine), which is a hormone produced in the pineal gland of the brain, has identified physiological properties that produce anti-inflam-matory and antioxidant effects. Although pineal gland is the primary source of MT production and release into the bloodstream, the receptors and enzymes responsible for the synthesis of MT are located in various tissues [9,10]. In addition, previous studies have identified MT as a direct free radical scavenger that neutralizes reactive oxygen species (ROS) and reactive nitrogen species (RNS). Moreover, MT has been shown to im-prove mitochondrial function by enhancing the activity of antioxidant enzymes, thereby preventing oxidative damage [11-13].
MT has recently been shown to provide several potential benefits including those of improved sleep quality and antioxidant regulation. These positive aspects support the potential of MT to supplement the recovery process of athletes [14]. This review summarized the recent evidence regarding the molecular structure and biological functions of MT and the potential role of this hormone in antioxidant and anti-inflam-matory processes.
METHODS
This study involved an extensive review of previous research using the PubMed, Science Direct, and Google Scholar databases. The keywords chosen for the searches were: “ melatonin” AND (“ antioxidant” OR “ an-ti-inflammatory” OR “ sleep”) AND (“ exercise” OR “ exercise training” OR “ high-intensity exercise” OR “ athletes” OR “ performance” OR “ intensive exercise”) AND (“ muscle” OR “ muscle strength” OR “ muscle power” OR “ muscle fatigue” OR “ oxidative stress” OR “ muscle damage” OR “ inflammation”).
Inclusion criteria followed was: 1) accessed to full-text articles, 2) published in English, 3) designed by randomized control study (RCT). Ex-clusion criteria followed was: 1) accessed by conference abstract and book, 2) published in other language than English.
RESULTS
1. Molecular structure and biological functions of melatonin
MT is a versatile and potent natural antioxidant that is produced by plants and mammals but can also be acquired through the consumption of fruits and vegetables [15]. The MT molecule is synthesized in specialized cells of the pineal gland called pinealocytes primarily during the dark phase of the day when there is a significant increase in the activity of the serotonin-N-acetyltransferase (arylalkylamine N-acetyltransferase; AA-NAT) enzyme. This enzyme converts 5-hydroxytryptamine (5HT; commonly known as serotonin) into N-acetylserotonin, which is further converted into MT through the action of the acetylserotonin O-methyl-transferase enzyme (Fig. 1) [16].

Molecular structure of melatonin.
MT is distinguished by its electron-rich aromatic indole heterocycle, which enables it to function as an electron donor and effectively reduce cell concentrations of electrophilic species. Furthermore, the side chains of MT present both hydrophilic and lipophilic properties, which allow for easy access across cell membranes, where the MT molecules can offer protection against oxidative damage [17,18]. In addition, the methoxy and amide groups present in MT enhance the free radical scavenging capabil-ity and complement the ability of the electron-rich indole to neutralize these harmful species. Furthermore, the electronic structure of the indole ring provides MT with high resonance stability, making it particularly ef-fective against ROS. This combination of structural elements reinforces the effectiveness of MT as both an antioxidant and ROS scavenger [19,20]. The carbon atoms (C-atoms) within the indole ring structure attract hydroxyl radicals (HO) and nitric oxide radicals (NO) due to their low ener-getic barrier, which facilitates adduct formation [14,21]. These properties allow MT to efficiently scavenge a wide range of reactive species, including peroxyl radicals (ROO), singlet oxygen, and peroxynitrite (ONOO-). In addition, MT inhibits nitric oxide synthase and hypochlorous acid (HClO) activities [22] and has the ability to transform into several metabolites using oxidative and enzymatic pathways. The resultant metabolites, including 3-OHMT, 6-OHMT, AMK, and AFMK, are potent free radical scavengers [14] and significantly contribute to the comprehensive antioxidant effects of MT. MT can be distinguished from commonly used antioxidants by its capacity to effectively neutralize up to ten categories of ROS/RNS. This characteristic establishes MT as a more potent antioxidant than conventionally used compounds [23].
2. Secretion of melatonin
The majority of organisms, including humans, display a physiological and behavioral pattern that follows a 24-hour cycle in harmony with their environment, which is referred to as a circadian rhythm. Circadian rhythms collectively form a circadian system that ensures the synchroni-zation of internal cellular clocks with daily environmental changes, especially the light-dark cycle [24]. The circadian system is hierarchically structured with a central pacemaker located in the suprachiasmatic nucleus (SCN) of the hypothalamus. The SCN serves as a master clock that regu-lates neuroendocrine functions, autonomic responses, and sleep-wake cy-cles [11]. At night, the SCN generates a neural output signal that triggers MT production in the pineal gland. Thus, exposure to light and engage-ment in physical activity regulate the SCN and suppress the production of MT (Fig. 2) [25,26].

Secretion of melatonin.
MT is produced through a series of biochemical steps. First, tryptophan is taken up from the bloodstream and converted into serotonin. Serotonin is then transformed into MT in a two-step process involving the sequential action of two enzymes: serotonin-N-acetyltransferase (NAT) and hydroxyindole-O-methyltransferase (HIOMT). Norepineph-rine then binds to adrenergic β1 receptors, which initiates the synthesis of MT. This binding activates pineal adenylate cyclase, which results in an increase in cyclic AMP (cAMP) levels and de novo synthesis of NAT or its activator. The inducible cAMP early repressor is a form of the cAMP-responsive element modulator that is activated in conjunction with NAT and limits the nighttime production of MT, contributing to the regulation of MT levels in the body [27,28].
MT is highly soluble in both lipids and water, which allows it to easily cross cell membranes, after which it is promptly released into the bloodstream rather than being stored within the pineal gland. Once established in the bloodstream, MT accesses various body fluids, tissues, and cellular compartments [29]. MT is primarily secreted during nighttime hours, with its highest levels in the bloodstream typically occurring at approxi-mately 03:00-04:00; however, the timing can vary based on the chrono-type of the individual. During the daytime, MT levels are either unde-tectable or relatively low, especially in well-rested individuals [27,30].
3. Role of melatonin: ant-inflammatory, antioxidant
MT is an immune modulator with pro- and anti-inflammatory properties, both in the context of diseases and in relation to mitochondrial function. It shows diverse effects on the immune system and mitochon-dria that suggest its therapeutic potential for a wide range of conditions. MT is a potent compound that improves mitochondrial function by acting as a robust antioxidant that scavenges ROS and RNS [31-33].
The reduction of ROS by MT occurs through two main mechanisms: direct free radical scavenging and antioxidant enzyme stimulation, particularly those of glutathione reductase, glutathione peroxidase, superoxide dismutase, and catalase. Specifically, MT stimulates the synthesis of intrinsic antioxidants that collaborate synergistically with other free radical scavengers. In addition, MT inhibits the activity of pro-oxidant en-zymes such as myeloperoxidase, eosinophil peroxidase, nitric oxide synthase, and cyclooxygenase-2 (COX-2), thereby indirectly affecting oxidative stress levels [34,35].
Mitochondrial dysfunction and inflammation are frequently associated with elevated nitric oxide (NO) levels due to effects induced by NO metabolites such as nitrosating derivatives (NO+, HNO, N2 O3, S-nitroso glutathione), peroxynitrite (ONOO−), and free radicals that arise from peroxynitrite [the hydroxyl radical (• OH), the carbonate radical (CO3•−), and nitrogen dioxide (• NO2)]. These RNS and oxidizing radicals can par-tially disrupt the electron transport chain (ETC) by binding to iron and iron-sulfur clusters, nitrosating and nitrating components, and oxidizing ETC elements [36,37]. Antioxidative protection and mitochondrial function maintenance mitigate the effects of inflammation by promoting an-tioxidative processes and reducing the formation of free radicals and excessive levels of NO. NO production is regulated through the suppression of inducible nitric oxide synthase (iNOS) and nitric oxide synthase (nNOS) activities, contributing to the control of inflammation [38,39].
MT has been reported to inhibit high-mobility group box-1 (HMGB1) signaling and toll-like receptor-4 (TLR-4) activation, which prevents the activation of the inflammasome NLRP3, inhibits the activation of nuclear factor kappa B (NF-κB) and upregulates the expression of nuclear factor erythroid 2-related factor 2 (Nrf2). In addition, the downregulation of proinflammatory cytokines and upregulation of anti-inflammatory cytokines aids in the control of inflammatory responses and the protection of cells from damage [40,41].
4. Effects of melatonin on exercise performance
Several factors influence athlete performance, including physical fit-ness, skill proficiency, nutrition, overall conditioning, and recovery. MT could enhance exercise performance by improving sleep quality, reducing oxidative stress, and supporting post-exercise recovery. Research investi-gating the influence of MT supplementation on physical performance is ongoing. Tables 1, 2, and 3 summarize 21 representative studies that have examined the effects of MT supplementation on exercise performance, focusing on sleep quality, antioxidant effects, and recovery processes.

Representative experimental findings on the role of melatonin in regulating sleep quality
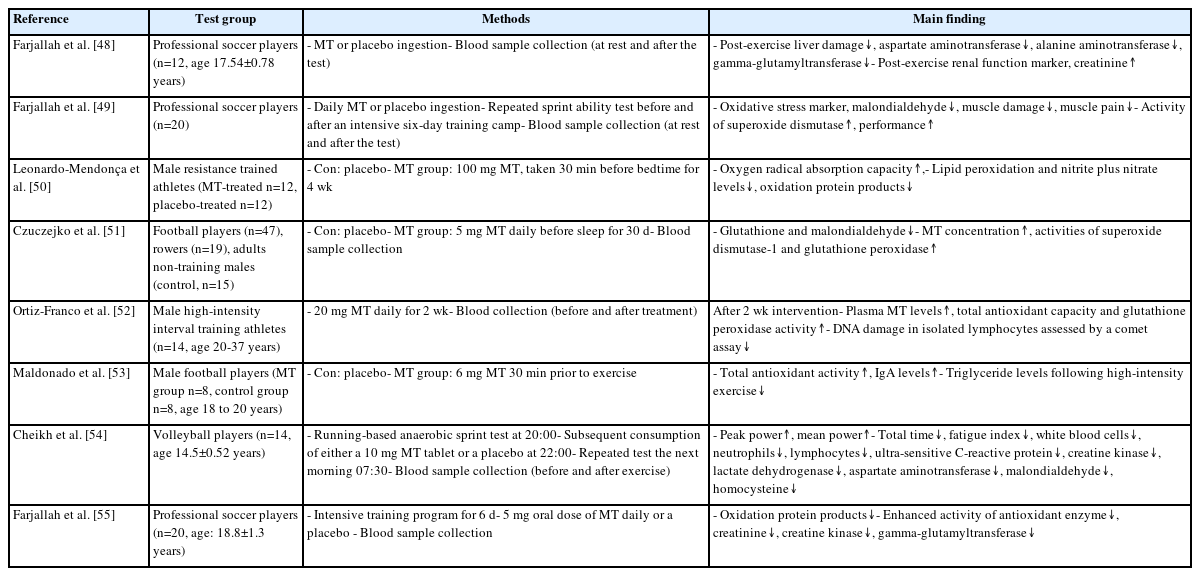
Representative experimental findings on the role of melatonin in antioxidation

Representative experimental findings on the role of melatonin in muscle damage
1) Sleep
Athletes must prioritize adequate and restorative sleep because the essential process of recovery occurs during this state. MT is closely associated with the quality of sleep and physical performance because when MT levels are elevated, tiredness and sympathetic nervous system suppression occurs, which ultimately contributes to high-quality sleep [42,43]. Table 1 shows representative studies that investigated the effects of MT on sleep quality, such as that of Oʼ Donnell et al. [44], who showed that a decrease in salivary MT levels was linked to total sleep duration following night training in elite female netball athletes. Cheikh et al. [45] provided evidence that the administration of 10 mg of exogenous MT after high-intensity evening training resulted in improved sleep quality, increased total sleep duration, and reduced sleep latency. In addition, Leonardo-Mendonça et al. [46] demonstrated the beneficial effects of MT in extending the duration of body temperature waveforms and maintaining a steady nocturnal state, which resulted in improved sleep efficiency in male resistance-trained athletes. Furthermore, Rostamdokht et al. [47] showed that MT supplementation effectively increased total sleep duration, decreased sleep initiation delays, and improved sleep overall quality in martial arts athletes who experienced dis-ruptions in their wake-sleep cycle and metabolic imbalances due to Rama-dan fasting. These findings suggest that MT is strongly associated with sleep quality and has the potential to enhance the recovery of athletes.
2) Antioxidants
Exercise provides sustained high-oxygen delivery to working muscles, including those involved in respiration. In elite athletes, this is largely achieved through an increase in cardiac output, adaptations of trained muscles, and amelioration of the deleterious effects of exercise-related oxi-dative stress. When exercise intensity reaches or exceeds 50% of the maxi-mum rate of oxygen consumption (VO2max), ROS/RNS levels are increased [14,24], and MT has a superior ability to efficiently neutralize ROS/RNS levels when compared to other antioxidant compounds [23]. Table 2 pres-ents an overview of representative studies that have explored the effects of MT on antioxidant activity. Farjallah et al. [48] demonstrated that MT intake resulted in a reduction of post-exercise indicators of liver injury (aspartate aminotransferase, alanine aminotransferase, and gamma-glutam-yltransferase) and an enhancement of post-exercise markers of kidney function such as creatinine. Oral MT supplementation has been shown to reduce oxidative stress (malondialdehyde) and muscle damage (creatine kinase and lactate dehydrogenase), enhance the activity of the primary antioxidant enzyme (superoxide dismutase), and alleviate muscle pain, thereby mitigating the performance declines that were observed in soccer players after training [49]. Leonardo-Mendonça et al. [50] demonstrated that supplementation with MT resulted in an increase in oxygen radical absorption capacity, a decrease in lipid peroxidation, and a reduction in nitrite and nitrate levels in resistance-training athletes. Czuczejko et al. [51] showed that MT supplementation caused a significant reduction in oxidative stress markers, specifically glutathione and malondialdehyde, in professional athletes who used intense training methods. These reductions were complemented by a significant increase in the MT concentration and the activities of superoxide dismutase-1 and glutathione peroxidase, which safeguarded tissues against the detrimental effects of reactive oxygen and reactive nitrogen species and inflammatory processes. Ortiz-Franco et al. [52] used a comet assay to reveal that the oral administration of MT resulted in increased plasma MT levels, which resulted in an improvement in total antioxidant capacity and a reduction in DNA damage in isolated lymphocytes. Maldonado et al. [53] showed that MT treatment in football players reversed oxidative stress and improved IgA levels and lipid metabolism. Moreover, Cheikh et al. [54] and Farjallah et al. [55] stated that MT intake reduced liver damage in both volleyball and soccer players. This reduction in liver damage was associated with increased antioxidant enzyme activity and decreased oxidative stress. These results suggest a strong con-nection between MT and antioxidant activity, indicating the potential of this hormone to enhance athletic performance.
3) Muscle damage
In addition to enhancing sleep quality, reducing oxidative stress, and improving post-exercise recovery, MT affects the rhythmic regulation of several physiological processes either directly or indirectly, which can enhance the physical performance of athletes. The ability of MT to read-ily cross the blood-brain barrier and enter subcellular compartments within cells contributes to its wide-ranging effects, including muscle recovery and anti-apoptotic and anti-inflammatory responses [56]. Table 3 summarizes the representative studies that examined the effects of MT on muscle damage.
Damaged muscle tissues undergo a muscle regeneration process that involves myolysis to remove excessive debris and break down non-functional muscle components. Muscle progenitor cells then activate, prolifer-ate, differentiate, and merge with existing myofibers to create newly restored functional myofibers [57,58]. MT promotes muscle recovery and hypertrophy by enhancing gene expression and increasing the number of satellite cells. Since these satellite cells are vital for muscle growth and regeneration, MT is a key factor for optimizing muscle health and performance. Su et al. [59] demonstrated a rapid increase in Pax7 expression that effectively restored skeletal muscle differentiation and improved muscle fiber morphology in mice with glycerol-induced muscle injury. MT is associated with muscle recovery and hypertrophy; therefore, MT levels are closely aligned with muscle strength and mass. Lee et al. [60] demonstrated a relationship between urinary MT levels and sarcopenia in postmenopausal women, and Obayashi et al. [61] found a correlation between urinary MT levels and muscle strength in patients with sarcopenia. Furthermore, Rondanelli et al. [62] revealed that administering MT in conjunction with essential amino acids increased total fat-free mass. These findings emphasize the impact of MT on muscle health and performance.
As previously mentioned, MT exerts neuroprotective effects by decreasing oxidative stress and inflammation and diminishing neural apoptosis through the activation of the Wnt/β-catenin signaling pathway [63,64]. A recent study by Mehanna et al. [58] demonstrated significantly increased twitch force and MT1a receptor mRNA levels in the injured soleus muscles of rats. In addition, those authors noted a decrease in the Bax level of the injured muscles, indicating the anti-apoptotic and anti-inflammatory actions of MT. Ma et al. [65] provided evidence that MT treatment significantly increased the cross-sectional area of muscle fibers in lambs. This effect was attributed to the ability of MT to elevate the growth hormone and testosterone levels, which mediated the apoptosis signaling pathways. Abdulwahab et al. [66] reported that the administration of MT led to significant normalization of serum glucose levels and improved insulin levels in diabetic rats. Moreover, MT effectively prevented oxidative stress and inhibited increases in proinflammatory cytokines and the expression of Bax, caspase-3, and P53, while enhancing anti-inflammatory cytokine IL-10 and anti-apoptotic protein Bcl-2. Although there is lack of supportive information for protective effects of MT on exercise-induced muscle damage to recommend to athletes, anti-inflammatory properties of MT could have beneficial performance-re-lated outcomes such as muscle recovery after intense workouts. A study conducted by Dhia et al. [67] demonstrated that MT intake mitigated the increase in inflammatory markers, including C-reactive protein and white blood cells, as well as oxidative stress and muscle damage in obese individuals after high-intensity interval exercise. Similarly, Ochoa et al. [68] indicated that MT prevented the overexpression of proinflammatory mediators induced by strenuous exercise in amateur athletes.
Consequently, MT has shown potential to enhance athletic performance through its anti-apoptotic and anti-inflammatory properties; therefore, it is a promising candidate for aiding in athlete recovery after intense physical activity. However, more research needs to be clarified the use of MT as a supplement for athletes.
CONCLUSIONS
MT has recently been recognized for its therapeutic effects and potential as a dietary supplement to support an exercise routine. This review focuses on the beneficial effects of MT, particularly those of improving sleep quality and mitigating exercise-induced oxidative stress and muscle damage. However, research on the use of MT as a supplement for athlete performance or recovery remains limited, and its effects based on factors such as timing and dosage have not been clearly established. Moreover, unpleasant side effects of melatonin have occasionally been reported. As melatonin's therapeutic significance is related to sleep, its diurnal ingestion could result in symptoms like sleepiness, alertness deficiency, poor coordination and loss of balance. Additionally, there have been observations of a slight increase in blood pressure linked to nitric oxide synthase downregulation [69]. Therefore, further research is required to investigate the specific mechanisms of MT and their positive and negative effects on performance in athletes.
Notes
The authors declare that they do not have conflict of interest.
AUTHOR CONTRIBUTIONS
Conceptualization: J Cho; Formal analysis: J Cho; Methodology: J Cho; Project administration: J Cho; Writing - original draft: J Cho, T Kim; Writing - review & editing: J Cho, T Kim.