서 론
비알코올성 지방간(non-alcoholic fatty liver disease)은 단순지방증(hepatic steatosis), 염증을 동반한 지방간염(steatohepatitis), 간경변 및 간경화(cirrhosis)를 포함하는 넓은 의미의 간질환(liver diseases)이다[1]. 비알코올성 지방간은 다양한 대사질환과 연관되어 간 내에 지방을 축적하는 특징을 가지고 있어 대사증후군의 간 내 발현(hepatic manifestation)으로 알려지고 있고, 심혈관질환으로 인한 사망(mortality)과도 밀접한 관련이 있는 것으로 보고된다[1,2]. 우리나라의 경우 서구화된 생활습관, 신체활동의 부족, 체력 저하 등이 원인이 되어 비알코올성 지방간의 유병률은 급격한 증가 추세에 있으며 일반성인에서 약 18%에 달한다고 보고하고 있다[3].
비알코올성 지방간은 병인학적으로 고지방 및 고열량 식이 결과로 인해 잉여 지방산(외인성)이 간으로 과도하게 유입되거나 혹은 비만 관련 인슐린 저항성으로 인해 지방조직의 중성지방 가수분해의 활성화로 인해 혈중으로 방출된 유리지방산(내인성)이 간으로 과도하게 유입됨에 따라 중성지방이 과도하게 축적되어 발병한다[4,5]. 간세포에서 유리지방산의 베타산화(β-oxidation)는 주로 퍼록시좀(peroxisome)에서 일어나며 다양한 조절인자들이 지질대사에 관여하는데 특히, PPAR-α (peroxisome proliferator-activated receptor alpha)는 유리지방산의 베타산화를 촉진한다[6]. 또한 최근 연구에 따르면 NAD+ 의존적 탈아세틸효소인 SIRT1(sirtuin1)은 PPAR-α 등의 전사인자들의 네트워크를 조절하여 베타산화를 통해 지방간 형성을 억제한다고 보고했다[7].
MicroRNA (miRNA)는 약 21-22개의 염기서열로 이루어진 작은 RNA molecules로 단백질 정보를 저장하고 있지는 않지만 다양한 유전자 발현을 전사 후 과정(post-transcriptional level)에서 조절한다[8]. miRNA는 전체 유전자의 60% 이상을 조절하는 것으로 알려졌는데 주로 세포 성장, 증식, 분화, 세포사멸 및 대사에 영향을 주는 것으로 알려졌다[9-11]. 최근 연구에 의하면 비알코올성 지방간이 지방간염 및 간경화로 진행되는 병리학적 과정에서 miRNA가 중요한 조절자 역할을 하는 것으로 알려진다[12-15]. 특히, SIRT1을 표적으로 하는 miR-34a는 마우스의 지방간뿐만 아니라 사람의 지방간에서도 발현이 증가된 것으로 나타나 지방간의 병리학적 진행과정에서 지질대사에 중요한 역할을 하는 것으로 보고된다[16,17].
한편, 규칙적인 운동은 비알코올성 지방간을 포함하여 당뇨, 비만, 고지혈증과 같은 다양한 퇴행성 질환을 예방하고 치료할 수 있는 비약물적인 수단으로써 안정성은 물론이고 효능과 효과가 입증되고 있다[18,19]. 특히, 규칙적인 운동을 통한 심폐체력의 향상은 심혈관질환 및 암으로 인한 사망률을 효과적으로 감소시키는 것으로 보고된다[20,21]. 그 예로 높은 심폐체력 수준과 비알코올성 지방간은 부적 상관관계가 있다고 임상연구를 통해 보고하였고 동물 연구에 따르면 고강도 운동은 중강도 운동에 비해 마우스 간의 중성지방의 양을 유의하게 감소시키는 것을 확인했는데 이는 고강도 운동이 중강도 운동에 비해 간의 아디포넥틴 수용체(adiponectin receptor 2) 발현을 증가시켜 베타산화를 증가시키고 염증성 반응을 완화시키는 것으로 확인하였다[22,23]. 따라서 비알코올성 지방간을 완화시키는 데 운동 트레이닝의 강도에 영향을 받을 것으로 판단되고 이때 miRNA의 발현 또한 강도에 영향을 받을 것으로 예상되지만 현재까지 보고된 바가 없다.
이러한 점에 착안하여 본 연구는 운동강도가 고지방식으로 유도된 비만 마우스의 지방간에 미치는 영향을 miRNAs와 그 표적 유전자 탐색을 통해 비알코올성 지방간에 대한 운동강도의 역할을 규명하는 것을 목적으로 하였다.
연구 방법
1. 실험동물
본 실험에 사용된 실험동물은 4주령된 수컷 C57BL/6 40마리를 오리엔트바이오(ORIENT Bio, Suwon, Korea)로부터 구입하여 1주간의 환경 적응 기간을 가진 후 케이지당 마우스 5마리씩 넣어 사육하고 물과 사료는 제한 없이 섭취하도록 하였다. 사육실 온도는 21±1℃로 유지하였고, 명기와 암기는 각각 12시간으로 조절하였다. 마우스의 체중은 매주 2회 측정하였다. 본 연구는 AAALAC International (Association for Assessment and Accreditation of Laboratory Animal Care International)인증기관인 S대학교 동물관리사용위원회(IACUC)의 검토와 승인을 거쳤으며, 실험동물의 관리와 사용에 관한 지침을 준수하며 시행하였다(과제번호 11-8).
2. 연구설계
총 40마리의 5주령 C57BL/6마우스를 무작위로 일반사료(standard chow, SC)를 먹는 집단(SC =10)과 60% 지방이 함유된 고지방식이(high-fat diet, HFD; D12492, Research Diet)를 먹는 집단(HFD=30)으로 나누어 8주간 비만을 유도하였다. 8주간 고지방식이로 비만이 유도된 마우스 30마리를 다시 비운동 집단(HFD, n=10), 고강도 인터벌 트레이닝 집단(HFD+INT, n=10) 그리고 중강도 트레이닝 집단(HFD+MOD, n =10)으로 구분한 후 고지방식이 처치와 운동처치를 15주간 병행하였다(Fig. 1).
3. 운동처치
운동은 마우스 트레이드 밀(Columbus Instruments, Inc., Columbus, OH)을 이용하여 총 15주간 주 5회의 고강도 인터벌 트레이닝과 중강도 트레이닝을 실시하였다. 고강도 인터벌 트레이닝은 경사도 5°에서 17 m/min 속도로 1분, 10 m/min 속도로 2분을 1 cycle로 하여 총 12회를 반복하였다. 중강도 트레이닝은 경사도 5°에서 10 m/min의 속도로 44분간 운동하도록 하였다. 고강도 인터벌 트레이닝 집단과 중강도 트레이닝 집단의 총 운동량(총 이동거리: 약 600 m)은 동일하게 설정하였다. 본 운동 시작 전과 후에 각각 8 m/min에서 10분간 준비운동과 정리운동을 실시하였다[23].
4. 측정변인
1) 혈액 및 간 조직 채취
실험이 종료된 실험동물은 12시간 절식시킨 후 마취시켜 간 조직을 채취하였다. 심장에서 채혈한 혈액은 원심분리기에 3,000 rpm에서 15분간 원심 분리하여 혈청을 분리하고 분석 전까지 -85℃에서 보관하였다. 적출한 간 조직은 조직면역학 분석을 위해 10% formaldehyde에 고정하여 분석 시까지 냉장 보관하였다.
2) 혈액변인 분석
혈청의 alanine aminotransferase (ALT)와 aspartate aminotransferase (AST) 수준은 Beckman DXC 800 analyzer (Brea, Ca)를 이용하여 측정하였고 total cholesterol (TC)는 enzymatic kits (Wako Chemicals USA, Inc., Richmond, VA)를 이용하여 측정하였다.
3) 조직면역학 분석
채취한 간 조직을 10% formaldehyde에 고정하고 파라핀으로 포매하여 5 μm 두께로 절편 슬라이드를 만들었다. 슬라이드를 흐르는 물에 헹군 뒤 haematoxylin (핵 염색)과 eosin (세포질 염색)을 사용하여 염색하고 봉입하였다. 조직표본은 광학현미경(Leica DMLS, Leica microsystems, Wetzlar, Germany)을 통해 관찰하고 사진촬영하였다.
4) Quantitative Real‐Time PCR
간 조직을 TRI reagent (Ambion, Austin, TX, USA)를 이용하여 균질화시킨 후, total RNA는 phenol 침전법을 이용하여 추출하였다. Total RNA의 농도는 Nanodrop (Thermo Fisher Scientific Inc., Rockford, IL, USA)을 사용하여 측정하였다. mRNA의 발현 정도를 확인하기 위해 30 ng의 RNA와 Taqman probe (PPAR-α: AB Assay IDs Mm00440939_m1, SIRT1: AB Assay IDs Mm01168521_m1) 및 TaqManTM RNA-to-CtTM1-Step Kit (Thermo Fisher Scientific Inc.)를 이용하였고, miRNA 발현은 추출된 RNA를 Taqman MicroRNA Reverse Transcription Kit (Thermo Fisher Scientific Inc.)를 이용하여 역전사한 후 Taqman MicroRNA Assay (Thermo Fisher Scientific Inc.)와 Taqman probe (miR-34a: AB Assay IDs 000426; miR-149: AB Assay IDs Mm04238124_s1)를 이용하여 Real-time PCR (Applied Biosystem, Foster City, CA, USA)로 정량화 하였다.
모든 PCR 반응은 3번 반복실험을 수행하였고, β-actin과 U6를 endogenous control로 이용하여 모든 표본의 발현을 균일화 하였다. mRNA 및 miRNA의 상대적 발현 수준은 2-ΔΔCt를 이용하여 계산하였다.
5) MicroRNA array 분석
추출된 total RNA는 Agilent Bioanalyzer® (Agilent Technologies, Santa clara, CA, USA)를 이용하여 RNA의 integrity를 확인하였다. RNA에 형광시료를 붙여, 799개의 mature microRNA에 상응하는 13,737개의 probes와 22개 control probes가 포함된 mouse miRNA microarrays (Agilent Technologies)에 hybridization을 시켰다. cDNA microarray는 50 ng의 RNA를 Aglient 44K oligonucleotide microarrays (Agilent Technologies)에 hybridization 시켰다. 각 칩에 control probes를 이용하여 보정 후 p-value가 .01 이하인 것을 선택하였다.
연구 결과
1. 운동강도가 마우스 체중 변화에 미치는 영향
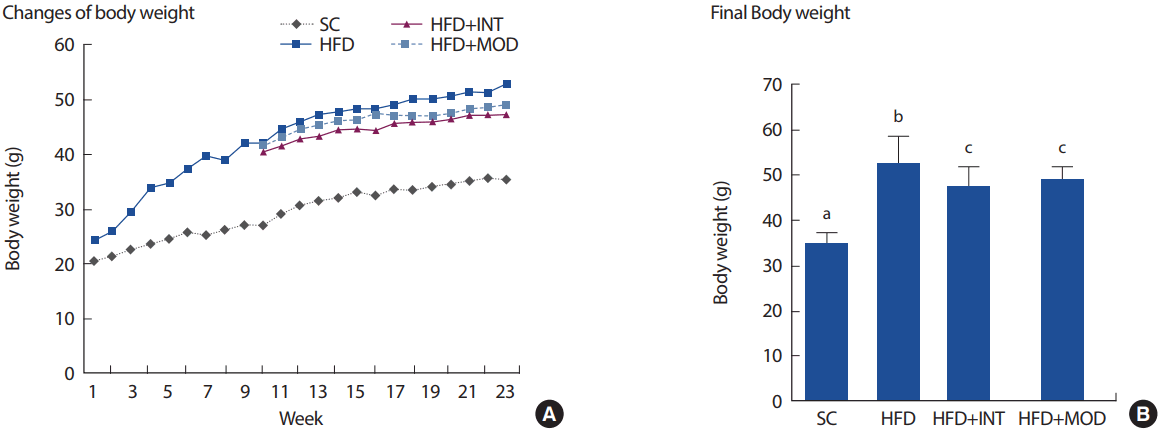
8주간 고지방식이 유도는 정상식이에 비해 체중이 약 1.5배 유의하게 증가하는 것으로 나타났다(SC: 27.0 ±1.5, HFD: 41.9±2.3, p <.001) (Fig. 2A). 8주간 고지방식이로 유도된 마우스를 비운동 집단(HFD)과 고강도 인터벌 트레이닝 집단(HFD+INT), 중강도 운동 집단(HFD+MOD)으로 나누어 15주간 고지방식이 처치와 동시에 운동트레이닝을 시킨 후 체중을 비교해 본 결과, 총 23주간의 고지방식이 처치는 정상식이 처치에 비해 유의한 체중 증가를 유도하였다(SC: 35.4±2.0, HFD: 52.8±6.2, p<.001). 그러나 15주간 운동 트레이닝은 고지방식으로 인한 체중증가를 억제하는 효과가 있었고 이때 운동 강도에 따른 유의한 차이는 나타나지 않았다(HFD+INT: 47.4±4.5, HFD+ MOD: 48.8±3.0) (Fig. 2B).
2. 운동강도가 간의 조직학적 특징, 간 지표 및 혈중 지질에 미치는 영향
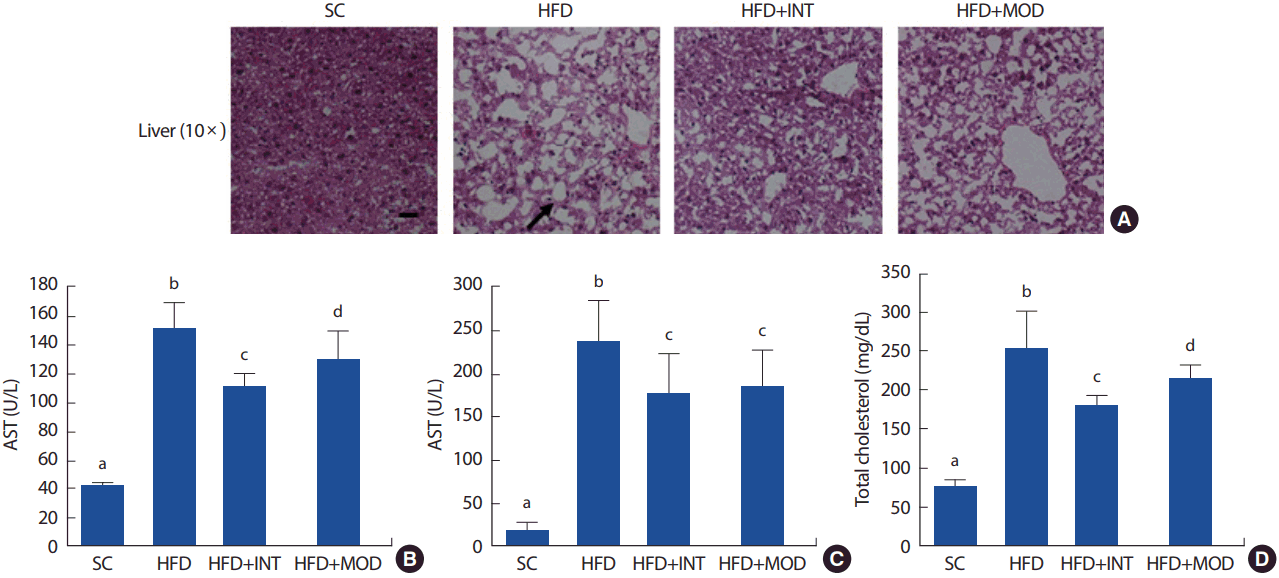
총 23주간의 실험 처치에 따른 간의 조직학적 변화 그리고 혈중 지질 및 간 지표에 대한 결과는 Fig. 3에 제시된 바와 같다. 간 조직 내 지방구의 형성은 정상식이에 비해 고지방식이 집단에서 지방입자의 수와 크기가 증가한 것으로 관찰되었다(Fig. 3A). 고지방식이는 정상식이에 비해 대표적인 간 손상 지표인 AST (SC: 41.6±2.1, HFD: 151.6±18.0, p <.001)와 ALT (SC: 19.4±7.6, HFD: 237.8±45.9, p <.001)의 양을 각각 3.6배와 12.2배 통계학적으로 유의하게 증가시키는 것으로 나타났고, 혈중 총 콜레스테롤 양을 약 3.3배(SC: 76.6 ±7.4, HFD: 253.4 ±48.5, p<.001) 유의하게 증가시키는 것으로 나타났다(Fig. 3B, C).
15주간의 운동트레이닝은 초기 8주간의 고지방식으로 인한 간 조직 내 지방구의 형성을 억제하는 것으로 나타났고, 이러한 운동 트레이닝의 효과는 중강도에 비해 고강도 인터벌 트레이닝 그룹에서 더 뚜렷한 것으로 나타났다(Fig. 3A). 뿐만 아니라 15주간의 운동트레이닝은 고지방식으로 인한 간 조직의 손상지표인 AST와 ALT의 증가를 억제시키는 것으로 나타났다. AST수준의 경우 중강도에 비해 고강도 인터벌 트레이닝 집단에서 억제효과가 더 뚜렷한 것으로 나타났고(HFD+INT: 110.5±9.5, HFD+MOD: 129.8±19.0, p<.05), ALT 수준의 경우 운동 강도에 따른 집단 간 차이는 없는 것으로 나타났다(Fig. 3B, C).
또한, 15주간 운동트레이닝은 혈중 총 콜레스테롤 양의 증가를 유의하게 억제시키는 것으로 나타났고, 이러한 운동 트레이닝의 효과 또한 중강도에 비해 고강도 인터벌 트레이닝 그룹(HFD+INT: 181.4 ±10.9, HFD+MOD: 215.3±163.4, p <.05)에서 억제효과가 더 뚜렷한 것으로 나타났다(Fig. 3D).
3. 운동강도가 간의 miRNA 발현 변화에 미치는 영향
고지방식으로 유도된 비만 마우스를 이용하여 운동강도에 따른 간 조직의 miRNA 발현량 변화를 확인하기 위해 적출된 간 조직의 miRNA array를 실시한 후, 23주간 고지방식이에 의해 dysregulation된 유전자를 운동트레이닝을 통해 통계학적으로 유의하게 완화시킨 유전자를 탐색하여 발굴하였다. 그 결과, 초기 8주간의 고지방식에 반응하여 간 조직의 miR-107, let-7d, miR-15b, miR-34a, miR-149의 발현이 증가되는 것으로 나타났고, miR-26a 발현은 감소되는 것으로 나타났다. 이에 반해 15주간 운동트레이닝은 간 조직의 miR-107, let-7d, miR-15b, miR-34a, miR-149의 발현의 증가를 유의하게 억제시키는 것으로 나타났고, miR-26a 발현의 감소를 완화시키는 것으로 나타났다(Table 1). 특히, 흥미로운 사실은 중강도에 비해 고강도 인터벌 트레이닝이 고지방식으로 인해 증가한 miR-149와 miR34-a 발현 증가를 효과적으로 억제시키는 결과를 유도했다는 점이다.
4. 운동강도에 따른 miRNA와 표적 mRNA의 발현 변화
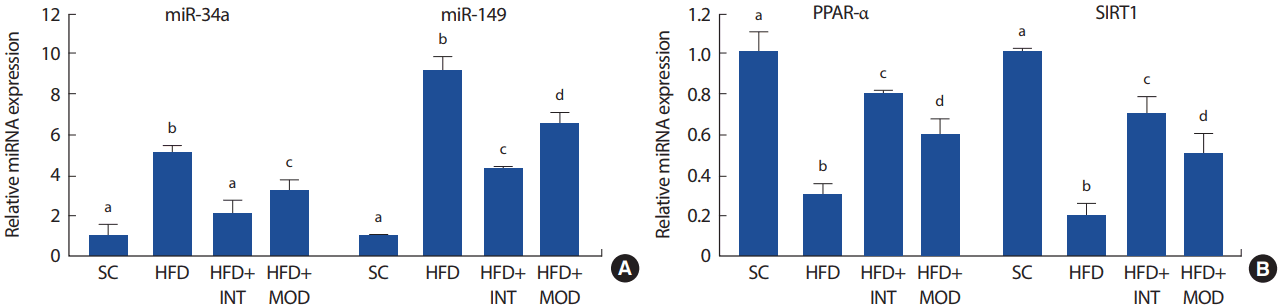
MicroRNA array 결과 고지방식이에 의해 가장 큰 변화를 보인 miR-34a와 miR-149를 real-time PCR로 정량화 하였다. 총 23주간 고지방식이 유도는 정상식이 집단에 비해 간 조직의 miR-34a와 miR-149의 발현량을 통계학적으로 유의하게 증가시키는 것으로 나타났다(Fig. 4A). 총 23주의 실험기간 중 후기 15주간의 운동트레이닝은 고지방식이로 인해 증가된 간 조직의 miR-34a와 miR-149의 발현량을 유의하게 억제하는 것으로 나타났고, 특히 고강도 인터벌 트레이닝 집단이 중강도 트레이닝 집단에 비해 뚜렷한 억제 효과를 나타냈다(HFD+INT vs HFD+MOD, p<.05).
이러한 사실을 고려하는 miR-34a의 표적 mRNA로 알려진 PPAR-α와 SIRT1의 유전자 발현을 정량화하였다. 그 결과 총 23주간 고지방식이는 정상식이에 비해 간 조직의 PPAR-α mRNA와 SIRT1 mRNA 발현량을 유의하게 감소시키는 것으로 나타났다(Fig. 4B). 그러나 23주 처치기간 중에 실시한 15주간의 운동트레이닝은 고지방식으로 인한 간 조직의 PPAR-α mRNA와 SIRT1 mRNA 발현량의 감소를 억제하는 효과가 있는 것으로 나타났고, 특히 고강도 인터벌 트레이닝 집단이 중강도 트레이닝 집단에 비해 상대적으로 더 뚜렷한 억제 효과가 있는 것으로 나타났다(HFD+INT vs HFD+MOD, p<.05).
논 의
본 연구는 고지방식으로 유도한 비만 마우스 모델의 지방간에 대한 운동 강도의 효과를 간 조직의 miRNA 및 표적 mRNA 유전자 발현에 미치는 영향을 중심으로 검증하는 것으로 주요 목적으로 하였다. 그 결과, 총 23주간 고지방식이 기간 중에서 실시한 15주간 운동 트레이닝은 마우스의 고지방식으로 인한 지방간을 완화시키는 효과가 있었고, 특히 운동 트레이닝에 따른 완화효과는 중강도에 비해 고강도 인터벌 트레이닝에서 더 명확한 것으로 나타났다. 흥미로운 사실은 고지방식이에 의해 비정상적으로 조절된 miRNA 중에서도 miR-149, miR-34a, miR-107, let-7d, miR-15b, miR-26a 발현량이 정상적인 방향으로 변화되었다는 것이고, 이 중에서도 특히 miR-149와 miR-34a는 고지방식이에 의해 가장 큰 변화를 보일 뿐만 아니라 운동 트레이닝에 의해서 가장 뚜렷한 변화를 보인다는 점이다. 또한. miR-34a의 표적 mRNA인 PPAR-α와 SIRT1의 발현을 조절하는 것으로 확인되었고, 비알코올성 지방간에 운동 트레이닝의 효과는 miRNA 조절을 통해 표적 mRNA를 조절되며, 그 효과가 운동강도에 따라 명확하게 차이가 있는 것으로 나타났다.
선행연구에 따르면 miR-34a는 단순지방간[16], 알코올성 지방간[24], 간경화[25], 간암[26] 등의 간 조직에서 발현이 증가될 뿐만 아니라 단순지방간 환자의 혈청[11]에서도 증가한다고 보고했다. Ding et al. [27]의 보고에 따르면 고지방식으로 유도된 비만 마우스의 지방간에서 miR-34a 발현이 증가되는 것을 확인하였고 이는 비알코올성 지방간이 진행될수록 점차 증가하는 선 경향을 보였다. 그러나 miR-34a 발현 억제제를 마우스에 주입했을 때 고지방식이로 유도했음에도 불구하고 간의 steatosis, 지방축적 및 혈중 AST의 수준 증가가 억제되는 것으로 나타났다. 특히 miR-34a는 PPAR-α와 SIRT1의 증가가 AMPK의 활성화를 증가시켜 지방산화를 높여 결국 지방간을 억제한다고 보고하였다. 이와 유사하게, Choi et al. [28]의 보고에 따르면 간에 miR-34a를 과발현 시켰을 때 PGC1-α와 PPAR-α가 감소된 반면 sterol regulatory element-binding protein-1 (SREBP-1)과 nuclear factor kappa B (NF-kB) 발현이 증가된 것으로 나타났다. 즉, 간에서 miR-34a의 과발현은 지방산화를 감소시키고 지방축적을 유도했을 뿐만 아니라 염증반응을 증가시켜 결과적으로 인슐린 저항성과 지방간을 유도하는 것으로 나타났다[16]. 본 연구 결과도 선행연구와 일치하게 23주간 고지방식이는 간 조직의 miR-34a의 발현을 증가시키는 것으로 나타났다. 즉, miR-34a의 발현 변화는 지방간의 병리학적 진행과정에 중요한 역할을 할 것으로 사료된다.
MiR-34a와 유사하게 miR-149도 고지방식이에 의해 간 조직에서 발현량이 증가되는 것으로 보고된다[29]. In vivo 실험을 통해 HepG2 세포에 miR-149 발현 억제제를 처치했을 때 지방산 합성이 억제되는 것으로 나타났고 이때 지방 대사에 조절자로 지방간 보호 효과를 주는 fibroblast growth factor-21 (FGF-21)의 발현을 증가시켜 간 세포의 지방 축적을 억제하였다[29]. 본 연구 결과도 총 23주간 고지방식이 유도는 간 조직의 miR-149의 발현량을 증가시키는 것으로 나타났다. 이는 miR-34a와 더불어 miR-149 또한 지방간의 병리학적 진행과정에 중요한 역할을 할 것으로 사료된다.
최근 연구에 따르면 운동은 간 조직의 miRNA를 조절한다고 보고하였다[30]. Xiao et al. [30]의 연구에 따르면 16주간 고지방식이를 유도했을 때 자발적인 휠 운동을 했던 집단에서는 지방간이 억제된 것을 확인하였고 간 조직에서 마이크로 어레이를 통해 분석해본 결과 고지방식이 그룹의 간 조직에서 miR-212가 과발현 되었으나 운동은 이를 완화시키는 것으로 나타났다. 운동에 의해 감소된 miR-212는 지방대사와 관련 있는 FGF-21의 발현을 증가시켜 지방간을 예방하는 것으로 나타났다. 또한 miR-149가 고지방식이로 유도된 지방간에서 발현량이 증가되었지만 16주간 자발적인 휠 운동에 의해 발현량의 증가가 억제되는 것으로 나타났다[30]. 고령의 당뇨병 환자를 대상으로 일회성 저항성 트레이닝을 실시했을 때 혈액의 백혈구에서 miR-146a가 인슐린 저항성 지표와 함께 감소되는 것으로 나타났다. 이처럼 운동은 miRNA의 발현 조절을 통해 표적 mRNA를 조절하여 다양한 세포 반응과 대사에 영향을 미치는 것으로 판단된다. 본 연구 결과에서도 고지방식이 유도로 인해 비정상으로 조절된 miRNA 발현이 운동을 통해 억제 혹은 완화되는 것을 확인하였고 특히, miR-34a와 miR-149 발현이 운동에 의해 가장 큰 완화효과를 보이는 것으로 나타났다. 또한 고강도 인터벌 트레이닝에서 더 큰 변화를 보이는 것으로 나타났으며 이는 운동 강도에 따라 간 조직의 miRNA의 발현이 달라질 수 있을 것이라 판단된다.
PPAR-α는 핵 호르몬 수용체로 포도당 항상성, 단백질 대사 및 세포 분화 및 지질대사에 핵심적인 역할을 하는 것으로 알려진다[31]. 특히, PPAR-α는 간에서 지방산화에 중요한 역할을 하는 것으로 알려졌는데 선행연구에 따르면 비만 쥐(OLETF)에 PPAR-α 길항제(agonist)를 처치했을 때 공복 시 혈당이 감소되고 지방간으로 진행이 억제되는 것으로 나타났으며 이때 간의 지방산화와 관련 있는 효소들의 작용이 활성화되는 것으로 나타났다[32]. 운동은 간의 PPAR-α 발현을 조절하는 것으로 보고되는데, Wang et al. [33]의 연구에 따르면 7주간 고지방식이로 유도된 비만 마우스를 중강도 운동 그룹과 고강도 운동 그룹으로 나누어 8주간 운동 트레이닝을 실시했을 때 간세포에 지방축적을 억제시키는 효과가 있는 것으로 관찰했고 특히 중강도 운동 그룹에 비해 고강도 운동 그룹에서 간 지방 합성과 관련된 유전자들이 감소되고 베타 산화와 관련된 PPAR-α와 같은 유전자의 발현이 증가되어 지방간 완화에 효과적인 것으로 보고했다. 또한 PPAR-α는 miR-34a의 표적 유전자로 보고되는데, Ding et al. [27] 연구에 따르면 간 세포에 miR-34a을 과발현시켰을 때 PPAR-α발현이 감소되는 것을 확인하였고 반대로 miR-34a의 발현을 억제했을 때 PPAR-α의 발현이 증가되는 것을 확인하였다. 본 연구에서도 23주간 고지방식에 따른 지방간의 PPAR-α 발현량은 정상식이 군에 비해 유의하게 감소된 것을 확인할 수 있었고 운동에 의해 miR-34a 발현의 감소와 더불어 PPAR-α 발현이 증가되는 것으로 나타났으며 운동강도에 의존적인 것으로 나타났다. 즉, 고강도 운동은 중강도 운동에 비해 간의 miR-34a 발현을 감소시키고 PPAR-α의 발현을 증가시켜 결과적으로 지방산화 활성화를 통해 지방간을 완화시키는 데 더 효과적이었을 것으로 사료된다.
SIRT1은 NAD+의존적인 탈아세틸 효소로서 NAD의 대사를 조절하고 세포의 대사를 조절하는 것으로 알려진다[34,35]. Choi et al. [28]의 연구에 의하면 간 조직의 miR-34a 발현 증가는 NAD(+)수준을 감소시키고 SIRT1 아세틸화를 활성화시켜 PGC1-α, NF-kB, SREBP-1c 발현 조절을 통해 비만을 유도하는 것으로 보고했다. 본 연구도 선행연구 결과와 유사하게 운동에 의해 증가된 간의 miR-34a는 SIRT1 발현을 증가시키는 것으로 나타났고, 이때 고강도 운동은 중강도 운동에 비해 SIRT1의 발현을 더 증가시키는 것으로 나타났다. 즉, 비알코올성 지방간에 운동은 miR-34a-SIRT1-PPAR-α의 신호전달 조절을 통해 지방간 완화에 긍정적인 효과를 나타내고, 중강도 운동에 비해 고강도 운동에서 그 효과가 더 큰 것으로 사료된다.
운동은 지방간 개선에 효과적이고 이때 운동 강도에 따른 효과가 있을 것으로 선행 연구에 의해 예상되고 있지만 명확한 하위 메커니즘이 밝혀져 있지 않은 실정에 본 연구는 운동 강도에 따른 간에서 miRNA 발현의 차이를 제시하였다. 그러나 추후 연구를 통해 운동 강도에 따라 다르게 발현하는 miRNA와 표적 mRNA 및 단백질에 대한 보다 광범위한 고찰이 필요할 것으로 사료된다.