장기간의 운동이 대장암 유발 쥐의 SPARC와 OSM발현에 미치는 영향
Abstract
PURPOSE:
The purpose of this study was to investigate the impacts of long-term treadmill exercise on the expression of SPARC and OSM in muscle, blood and colon polyp of colon cancer-induced mice.
METHODS:
Thirty mice were randomly divided into two groups: Exercise group (EX, n=15), Control group (CON, n=15). Mice of EX, at 6-week of old, performed exercise at moderate intensity for 30 minutes, 5 days per week, during 12 weeks. At 17-week of old, all mice were injected AOM and DSS to induce colon cancer. Then EX kept doing exercise for another 12 weeks. The expression of OSM and SPARC in muscle, blood, colon polyp was analyzed using the ELISA kit.
RESULTS:
The OSM expression of EX was significantly higher than CON in muscle (p<.01), but not in blood and polyps. On the other hand, the SPARC expression of EX was significantly increased in colon polyp (p<.01) compared with CON, however, there is no significant difference in blood and muscle.
CONCLUSIONS:
A 24-week treadmill exercise induced over-expression of SPARC and OSM in different tissues of colon cancer-induced mice, therefore these could be one of important factors to demonstrate exercise effect on cancer management.
색인어: 대장암, 트레드밀운동, SPARC, OSM, 마이오카인
Keywords: Colon cancer, Treadmill exercise, SPARC, OSM, Myokine
서론
마이오카인(myokine)은 근육에서 생성되어 분비되는 사이토카인(cytokine)을 말하며[ 1], 사이토카인의 역할과 마찬가지로 자가분비(autocrine)하여 근육의 성장과 대사를 조절하는 것은 물론, 인접한 세포에 작용하거나(paracrine), 순환하여 다른 기관의 표적 세포에 작용하여(autocrine) 세포간 활동을 조절한다[ 2]. 때문에 인체 기관 중 넓은 분포를 차지하고 있는 근육계는 마이오카인의 생성 및 분비로 인해 내분비 기관의 역할까지 담당할 중요한 기관으로 여겨지고 있다[ 1]. 또한 마이오카인은 안정시보다 근육의 수축활동 시 더욱 과발현이 되는 것으로 보고되면서, 신체 활동과 운동 역시 마이오카인의 생성에 중요한 역할을 하는 것으로 기대되고 있다[ 3]. 특히 운동으로 인해 증가된 혈중 사이토카인 중 마이오카인이 차지하는 비율이 지방 조직에서 분비되는 사이토카인의 양보다 많은 것으로 나타나고 있으며[ 4], Interlukin-6 (IL-6), IL-10, IL-15, Tumor necrosis factor-α (TNF-α) 등이 운동유발성 마이오카인으로 잘 알려져 있다[ 5]. 최근 연구들에서는 일부 마이오카인이 암을 예방하고 치료하는 데 중요한 기전을 담당하는 것으로 밝혀졌다. Secreted Protein Acidic and Rich in Cysteine (SPARC)은 사이토카인 중 하나로 세포와 기질 간 상호작용과 세포기능을 유도하는 작용을 하는 세포기질(matricellular) 단백이다[ 6]. SPARC는 Platelet-derived growth factor (PDGF), Vascular endothelial growth factor (VEGF) 등과 같은 성장 인자들을 통해 세포의 형태변화와 증식에 관여하는데[ 7, 8] 부적절한 SPARC의 발현은 암의 발생과도 밀접하게 연관되어 있다[ 9]. 암 종에 따라 양상이 다르기는 하나 유방암, 난소암, 췌장암 등에서 SPARC의 발현 감소를 보고하였고[ 10], 이처럼 종양 내 발현 감소나 비활성화된 경우 암 전이의 촉진이 유도되며, 반대로 활성화나 발현이 증가할 경우 암 전이가 저하된다고 하였다[ 11]. 특히 대장암의 경우 그 발현량이 현저히 감소되어 있는데 이는 유전자 촉진부(promoter) 과메틸화(hypermethylaton)로 인해 억제되었으며[ 12], 세포자멸(apoptosis) 단백질인 caspase-3, caspase-8 등의 조절이상과 연관되어 암 세포의 증식억제와 사멸에 부정적인 영향을 끼친다[ 13]. 따라서 SPARC의 발현은 대장암의 진행이나 관리에 있어서 매우 중요하며, 이에 따라 SPARC의 조절을 통한 화학적 치료가 암환자의 치료를 위해 사용되고 있다[ 14]. 이러한 최근 SPARC가 운동에 의해 인체 내 발현이 증가하는 것이 알려지고, 암의 관리에 있어서도 긍정적인 가능성을 보고하고 있다. Aoi et al. [ 13]은 건강상태가 양호한 일반인을 대상으로 한 시간의 일회성 사이클 운동을 실시한 결과 운동 직후 SPARC의 발현이 유의하게 증가하였고, 같은 대상자들을 4주간의 트레이닝한 이후에도 발현량이 유의하게 증가한 것을 보고하였다. 또한 Azoxymethane (AOM)으로 대장암이 유발된 쥐의 경우 6주간 저강도의 운동에 의해 근육에서의 SPARC mRNA가 유의하게 향상되었음을 보고하였으며, 특히 대장 조직에서 아무런 변화가 나타나지 않은 것과는 달리 근육-특이적으로 발생했음을 보고하였고[ 13], 다시 말해 SPARC 발현 증가를 위한 골격근 사용의 중요성을 입증하고 있는 결과로 여겨지고 있다. 암 세포자멸과 관련이 있는 또다른 마이오카인으로는 IL-6 family cytokine로 알려진 Oncostatin M (OSM)이 있다. OSM은 다양한 조직에서 다면 발현이 되며 염증 조절에 관여하는 것은 물론 caspase 활성을 통해 종양 세포의 증식을 억제하는 것으로 알려져 있다[ 15, 16]. 이러한 OSM 역시 운동 자극에 의해 근육에서 과발현이 되는 것으로 알려져 있어[ 5] SPARC와 함께 운동의 효과성을 설명하는 중요한 기전으로 기대되고 있다. 일반적으로 사이토카인들은 운동의 형태(type)와 운동강도(intensity)에 의해 다른 발현 양상을 보이는 것으로 알려져 있다. IL-6, IL-11, OSM, Leukemia Inhibitory Factor (LIF) 등은 운동직후 안정 시의 100배 이상 과발현되는 것으로 알려져 있으며[ 2], 운동의 강도가 증가함에 따라 발현량이 더욱 증가하는 것으로 알려져 있다[ 17]. 운동 종류에 따라서 발현 종류와 양상도 달라지며 혈관신생과 관련 있는 IL-8, 근육 성장의 동화작용을 일으키는 IL-15 등은 저항성 운동에 의해 과발현이 되는 것으로 알려져 있다[ 18]. 따라서 운동 강도나 형태는 이러한 사이토카인 혹은 마이오카인의 발현량과 밀접한 연관이 있는 것으로 여겨지고 있다. 그러나 OSM과 SPARC의 발현을 살펴본 선행연구들은 대부분 일회성 운동이거나[ 19, 20], 자발적 형태의 운동(voluntary wheel running)으로, 규칙적 운동의 효과를 검증하기 위한 지구성 트레이닝을 실시한 연구는 미흡한 실정이다. 또한 암 모델에서 운동에 의해 OSM과 SPARC의 발현성을 확인한 연구는 매우 미흡한 실정이며, 운동의 강도나 장기간의 운동이 미치는 영향을 살펴본 연구도 거의 전무한 실정이다. 그러므로 본 연구에서는 24주간의 장기간 트레드밀 운동을 실시하여, 종양세포의 증식과 사멸에 관련이 있다고 알려진 SPARC와 OSM 발현을 살펴보고, 이를 통해 암에 대한 운동의 효과성을 입증하기 위한 기초자료로 제시하고자 한다.
연구 방법
1. 연구 대상
본 연구에서는 출생시기(4주령, 18 g)가 동일한 Imprinting Control Region (ICR) 수컷 쥐(n =30)를 사용하였으며, 통제군(Control, CON), 운동군(Exercise, EX)으로 15마리씩 무선 배정하였다.
모든 쥐들은 S 대학교 G 연구소 내 동물 실험실에서 명암 주기가 12시간 단위로 바뀌고(08:00-20:00), 20˚C±2˚C로 온도가 유지되는 환경에서 사육되었다. 또한 물과 사료는 자유롭게 제공되었다.
2. 운동 프로토콜
본 연구에서는 장기간 운동의 예방적 효과가 암 모델에 미치는 영향을 검증하기 위해, 24주간의 유산소 운동을 실시하도록 하였으며 운동 중재 중 대장암을 유발하도록 하였다. 이는 본 연구에서 적용한 대장암 유발 방법이 유발 시작 시점부터 최종적으로 암 유발을 확인하는 데까지 최소 80일(약 10주)의 기간을 필요로 하므로[ 21], 해당 기간을 포함하여 운동 기간을 설정함으로써 성공적인 암 유발 확인과 함께 장기간 운동의 효과를 적용하기 위함이다( Fig. 1). 본 운동프로그램은 다음과 같다. 6주령의 쥐를 대상으로 준비운동과 정리운동을 포함하여, 일일 30분, 주 5회, 총 24주간 점증적 부하의 유산소 트레드밀 러닝을 실시하였다. 운동 프로그램의 강도는 Bedford [ 22]가 사용한 실험 쥐를 위한 트레드밀 운동 강도와 암 모델에서 적용해 온 운동 강도를 참고하여 중강도(60-75% VO 2max)로 설정하였으며, 이에 따라 1주차에 14 m/min으로 시작하여 2주마다 1 m/min씩 점증 부하 하였고, 운동 시작 9주에는 18 m/min가 되게 하였다. 또한 대장암 유발을 시작한 시점(운동 12주차)부터는 강도 증가에 대한 스트레스를 줄이기 위하여 속도를 유지하도록 하여, 24주차까지 최종 트레드밀 속도는 18 m/min가 되었다. 또한 암세포에 영향을 미칠 수 있으므로 전기 자극은 사용하지 않았으며, 통제군은 운동군과 같은 환경에 노출시키기 위해 운동을 실시하는 동안 트레드밀 옆에 배치하였다. 구체적인 운동프로토콜은 Table 1과 같다.
3. 대장암 유발
본 연구에서는 대장암 유발을 하기 위해 Azoxymethane (AOM)과 Dextran Sodium Sulfate (DSS)를 병행하여 사용하는 방법을 이용하였다. AOM은 DNA를 알킬화하는 방법으로 암을 유발하는 화학제이며, DSS는 대장염을 통해 암을 유발하는 화학제이다. 본 연구에서 사용한 AOM/DSS 병행 방법은 만성 염증(chronic inflammation)에 의해 대장암에 걸리게 되는 방법으로, 80일 정도 후 대장암 유발을 성공적으로 확인할 수 있는 방법으로 알려져 있다[ 21]. 구체적인 대장암 유발 방법은 다음과 같다. 17주령의 쥐의 복강에 체중당 10 mg/kg의 AOM을 1회 주사하였고, 17, 20, 23주령에 DSS를 식염수에 2.5% wt/vol로 희석하여 제공하였다. 29주령에 일부 쥐들을 희생하였으며, 폴립의 생성을 통해 대장암 유발을 확인하였다. 30주령에 분석을 위해 모든 쥐들을 희생시켰으며, 대장암을 유발한 모든 쥐들에게서 폴립의 생성을 확인할 수 있었다.
4. 측정항목 및 방법 자료처리방법
1) 조직 적출 및 혈액 채취
24주간의 유산소 운동을 종료 후, 12시간 이상의 공복 후 안정을 취한 후 이산화탄소를 이용하여 마취시켰다. 체중을 측정한 뒤 혈액을 채취하고 대장 조직을 적출하였으며, 대장을 3부분으로 자른 다음 폴립 유무를 확인하였다. 근육은 비복근(Gastrocnemius)을 기시점과 부착점에 가깝게 절단한 후 정밀계량기(Sartorius BP61S, Germany)를 이용하여 무게를 측정하였다. 절취한 골격근과 폴립은 0.9% NaCl에 0.16 mg/mL의 헤파린이 첨가된 용매에 담아 적혈구를 제거한 후 homogenizer를 이용하여 4˚C에서 균질화(homogenization)한 후 원심분리하여 상층액을 제거하였다(3,000×g, 4˚C, 10분). 모든 조직 샘플은 Tissuetek으로 마운팅(mounting)을 한 후 액화질소에 넣어 급속 냉동시킨 후 분석할 때까지 초저온 냉동고(-70˚C)에서 보관하였다.
2) 체중 측정
동물 집단별 체중의 변화는 전자저울(mettler instrument AG CH-8606, Switzerland)을 이용하여 2주에 1회씩 측정하였다. 또 복부지방과 골격근 중 비복근(Gastrocnemius)을 최종 샘플 채취 시 최대한 기시점(origin)과 부착점(insertion)에 가깝게 절단하여 정밀계량기(Sartorius BP61S, Germany)를 이용하여 계량하였다.
3) Myokine 분석
혈액, 근육, 대장 폴립에서 SPARC와 OSM의 정량적 분석을 위해 효소결합 면역흡수 분석법(Enzyme-linked immunosorbent assay, ELISA) kit (Uscn life science, USA)를 절차에 따라 사용하였다. 모든 시약과 샘플을 준비하고, 각각의 웰(well)에 100 µL의 sample과 standard를 분주한 후 37˚C에서 두 시간 배양하였다. 내용물을 흡입하고 Detection Reagent A를 100 µL씩 분주한 후 37˚C에서 1시간 배양하였다. 내용물을 버리고 세척(wash)하기를 3번 반복하였다. Detection Reagent B를 100 µL을 분주한 후 잘 혼합하여 37˚C에서 30분 배양한 후 세척하기를 5번 반복하였다 90 µL 기질액(Substrate Solution)을 분주하고 다시 37˚C에서 15-25분 배양하였다. 각각의 웰에 반응정지액(Substrate Solution)을 50 µL씩 분주하고 Micro plate reader를 이용하여 흡광도 450 nm 필터에서 측정한 후 standard curve를 이용하여 농도를 산출하였다.
5. 자료처리방법
본 연구에서 얻어진 모든 자료는 분석을 위해 윈도우용 SPSS 통계프로그램(ver.21.0)을 사용하였다. 체중과 근육량은 평균(M)과 표준편차(SD)로, SPARC와 OSM은 평균(M)과 표본오차(SEM)로 나타내었다. 집단 간 차이를 검증하기 위해 독립표본 t-검정을 실시하였으며, 유의수준은 p< .05로 설정하였다.
연구 결과
1. 장기간의 트레드밀 운동이 대장암 유발 쥐의 체중변화에 미치는 영향
본 연구의 기간 동안 측정된 실험 쥐들의 체중 변화는 Fig. 2와 같다. 17주령과 30주령의 시기를 제외한 모든 주령에서 그룹 간 통계적으로 유의한 차이가 나타나지 않았다( p>.05). 17주령은 대장암 유발을 위해 AOM을 주입한 시기로 운동군(EX)의 체중이 통제군(CON)에 비해 유의하게 낮은 것으로 나타났으며( p < .05), 30주령은 AOM/DSS 방법에 의해 대장암의 유발을 확인할 수 있는 시기(AOM 주입 후 10주 이상)로, 운동군(EX)의 체중이 통제군(CON)에 비해 유의하게 높은 것으로 나타났다( p< .05).
2. 장기간의 트레드밀 운동이 대장암 유발 쥐의 신체조성에 미치는 영향
본 연구에서 대장암 유발 쥐의 근육량과 지방량을 측정한 결과는 Table 2와 같다. 24주간의 트레드밀 운동에 참여하였던 대장암 유발 쥐의 비복근(gastocnemius)량과 대퇴사두근(quadriceps)량은 통제군(CON)과의 비교에서 유의한 차이를 보이지 않았다( p>.05). 그러나 24주간의 트레드밀 운동에 참여하였던 대장암 유발 쥐의 복부 내장 지방(abdominal fat)량은 통제군(CON)에 비해 유의하게 낮게 나타남을 관찰하였다(p< .05).
3. 장기간의 트레드밀 운동이 대장암 유발 쥐의 SPARC에 미치는 영향
장기간 운동에 의해 대장암 유발 쥐의 근육, 혈액, 대장 폴립에서의 SPARC 발현을 분석한 결과는 Fig. 3에 나타난 바와 같다. 24주간의 트레드밀 운동에 참여하였던 운동군(EX)의 SPARC 발현은 통제군(CON)에 비해 대장 폴립에서 유의하게 높은 것으로 나타났다( p < .01). 그러나 비복근(gastrocnemius)과 혈중(blood)에서는 집단 간 유의한 차이가 없는 것( p>.05)으로 나타났다( Fig. 3).
4. 장기간의 트레드밀 운동이 대장암 유발 쥐의 OSM 발현에 미치는 영향
24주간의 트레드밀 운동에 참여하였던 운동군(EX)의 OSM 발현은 통제군(CON)에 비해 비복근(gastrocnemius)에서 유의하게 높은 것으로 나타났다( p< .01). 그러나 혈중(blood)과 대장 폴립(polyp)에서는 집단 간 유의한 차이가 없는 것( p>.05)으로 나타났다( Fig. 4).
논의
1. 장기간의 트레드밀 운동이 대장암 유발 쥐의 체중 변화에 미치는 영향
최근 많은 연구들이 체중이 5-10%라도 감소되었을 때 신체적으로 긍정적인 효과를 가져올 수 있다고 하였지만[ 23] 부적절한 체중감소 역시 부작용과 문제점을 초래한다. 특히 종양세포의 대사과정에서 LMF/ZAG, TNF-α, IL-6, IFN-γ 등과 같은 염증성 사이토카인의 증가에 따라 근육과 지방의 급격한 감소로 체중이 감소하는 현상이 나타나는데, 암 환자의 경우 5%의 체중감소는 삶의 질을 감소시키고 항암치료의 부작용과 더불어 결과적으로 생존율을 감소시킨다고 하였다[ 24- 26]. 본 연구에서 17주령부터 AOM/DSS 방법에 의해 대장암을 유발시켰다. 운동군(EX)의 경우 AOM을 주입하고 DSS를 음용한 17주령에 체중이 4주전에 비해 약 12% 정도 감소하였고, 통제군(CON)과 비교하여 현저하게 체중이 감소한 것을 확인할 수 있었다( p=.013). 이는 운동군(EX)에서 통제군(CON)과 동일하게 AOM/DSS를 통해 암 유발을 실시하였으나 운동 역시 스트레스를 유발하는 요인으로, 운동과 병행하여 암을 유발함으로써 스트레스가 가중되어, 체중의 유의한 감소를 가져온 것으로 사료된다. 또한 본 연구에서 실시한 암 유발방법은 AOM 주입 이후 10주가 지난 시점부터 대장암 유발을 확인할 수 있는 방법으로 27주령 이후부터 통제군(CON)의 체중이 감소하는 것을 확인할 수 있었으며 30주령째 통제군(CON)의 체중은 4주 전(27주령째)에 비해 약 11% 정도 감소하였고, 같은 주령의 운동군(EX)와 비교하여도 유의하게 낮은 것을 확인할 수 있었다( p=.026). 대장암 환자는 본래 체중의 약 30%까지 감소하고, 이러한 체중의 감소는 대장암으로 인한 사망 원인의 약 20-33%를 차지하는 것으로 알려져 있다[ 27]. 운동은 SOD 등과 같은 항산화 방어 기전의 증가와 NK 세포, IL-2 등의 항종양 면역방어체계를 촉진시켜 암에 대한 위험을 줄이는 것으로 알려져 있으므로[ 28, 29], 본 연구에서 장기간의 규칙적인 운동은 암이 유발된 이후 항염증 사이토카인 등으로 인해 발생할 수 있는 체중 감소 현상을 억제하는 역할을 한 것으로 사료된다.
2. 장기간의 트레드밀 운동이 대장암 유발 쥐의 신체조성에 미치는 영향
대장암 환자에게 있어 체중 유지와 함께 중요하게 고려되고 있는 것은 바로 근육량 유지이다. 대장암 환자에게 나타나는 근 감소현상은 암 환자들의 만성 염증, 피로, 대사 장애 등과 같은 증상들과 밀접한 연관이 있는 것으로 알려져 있으며[ 30, 31], 근육량 감소와 근 기능 감소는 호흡 관련 근육 기능 저하를 일으키고 사망에 까지 이르게 하는 것으로 알려져 있다[ 27]. 특히 암 악액질(cancer cachexia) 증상이 나타난 암 환자의 경우 근육량을 유지하는 것은 암환자의 생존율을 높이는 것으로 알려져 있어[ 32], 근육량의 유지 및 향상을 위한 운동 중재는 암의 진행과 치료에 있어서 중요한 요인으로 자리잡고 있다. 그러나 본 연구에서 운동군(EX)의 근육량은 통제군(CON)과 유의한 차이를 나타내지 못하였다. 이러한 결과는 지속적인 운동에도 불구하고 암 유발쥐의 근육량에 유의한 변화를 가져오지 못한 Daneryd et al. [ 33]의 결과와 일치하며, AOM/DSS 방법에 의해 대장암을 먼저 유발한 뒤 12주간의 운동을 실시하였으나 운동군과 통제군의 근육량에 유의한 차이를 가져 오지 못한 선행연구의 결과와 일치한다. 일부 대장암 모델(APC+) 쥐의 경우 운동에 의해 근육량이 증가하였음을 보고하였으나[ 34, 35] 이와 같은 상이한 결과는 대장암 유발 방식의 차이로 여겨진다. APC+ 쥐의 경우 근육 내 세포자멸을 조절하는 단백질(bax, bcl-2 등)의 발현 이상으로 근육감소는 물론 지방의 현저한 감소로 암 악액질(cancer cachexia) 현상이 나타나는 것으로 알려져 있다[ 36]. 비록 본 연구에서는 근육세포 자멸과 관련된 기전에 대해 분석하지 않았으나, 이러한 기전이 AOM에 의해 유도된 암 모델이나 운동의 중재에 따라 유의미하게 변화하지 않은 것으로 사료되며, 그로 인해 근육세포나 근육량에 영향을 미치지 않은 것으로 보인다. 이에 대해 관련 기전에 대한 연구가 추후 필요할 것으로 사료된다. 그러나 본 연구에서는 운동군(EX)의 복부 지방량이 통제군(EX)에 비해 약 3배가량 감소한 것( p=.014)으로 나타났다. 대장암 모델에서 복부지방의 감소는 매우 의미가 있다. 특히 비만은 대장암의 위험을 증가시키는 것으로 알려져 있으며[ 37, 38], 허리-엉덩이 둘레비(WHR)는 성인 여자의 대장암 용종 발생과 밀접한 연관이 있는 것으로 알려져 있다[ 39]. 이러한 기전에는 IL-6, TNF-α 등의 염증성 사이토카인의 혈장 농도 증가[ 40]와 혈장 인슐린 수치 증가에 따른 IGF-1과의 연관성과 관련이 있으며[ 41], 운동은 이러한 염증성 사이토카인의 분비 억제[ 42]와 혈중 인슐린 농도 등을 조절하는 것으로 지방 증가를 억제시키는 기전을 작용한다[ 43]. 선행 연구들의 경우 대장암 환자를 대상으로 저항성 운동을 실시하여 체지방, 몸통지방의 감소를 보고하였으며[ 44], AOM/DSS에 유도된 쥐모델에서도 자발적인 러닝에 의해 지방의 감소가 유의하게 나타났음을 보고하며[ 45] 본 연구 결과와 일치함을 보여주었다. 따라서 장기간 트레드밀 운동은 AOM/DSS로 유도된 대장암 쥐의 복부 지방 감소에 효과적으로 작용한 것으로 여겨진다.
3. 장기간의 트레드밀 운동이 대장암 유발 쥐의 SPARC 발현에 미치는 영향
SPARC는 세포 부착(cell adhesion), 혈관형성(angiogenesis), 세포 분화(differentiation) 등의 역할을 하는 사이토카인으로[ 46], 인체의 대장암 발생 또는 악화와 관련하여 그 발현이 현저하게 감소되어 있어, 대장암 환자에 있어서 SPARC의 발현 증가는 긍정적인 예후를 위한 중요한 요인이 되고 있다[ 47]. 본 연구에서 총 24주간의 중강도 트레드밀 운동에 참여한 운동군(EX)의 대장 폴립에서 SPARC의 발현이 유의하게 높게 나타났다( p =.001). 또한 통계적으로 유의하지는 않았으나 운동군(EX)의 비복근(gastrocnemius)에서 통제군(CON)에 비해 상당한 발현을 나타내었고( p =.055), 혈중에서는 통계적으로 두 그룹 간 유의미한 차이가 나타나지 않았다( p=.079). 이와 같은 결과는 운동에 의해 SPARC의 발현이 증가하였다는 선행연구와 일치하나 발현 조직에 있어서는 차이가 나타난다. Aoi et al. [ 13]은 정상 쥐를 대상으로 1회성 운동에 의해 간, 지방 조직, 대장보다 근육에서 특이적인 발현이 나타난다고 보고하였으며[ 13], 정상 성인의 1회성 사이클 운동은 운동 직후 혈액에서 유의한 증가를 보이나 6시간 이내 안정 시 수준으로 복귀하였음을 보고하였고, 같은 대상자로 4주간의 지속적인 운동 후에도 안정 시 혈중 SPARC 수준은 변함이 없고, 운동 직후 유의한 증가가 나타났다고 보고하였다. 따라서 24주간의 마지막 운동이 끝나고 하루가 지난 후 채혈한 본 연구의 혈중 SPARC는 운동 후 안정시 수준으로의 복귀로 통제군과 차이가 나타나지 않은 것으로 여겨진다. 그러나 폴립에서의 유의한 증가와 근육에서의 과발현 경향성은 운동의 효과로 여겨진다. 암과 관련하여 SPARC는 IL-8, Leukotrienes, Fas/FasL, IL-1 등과 같은 염증반응 인자들과 관련이 있고[ 48], 감소된 SPARC는 종양세포의 증식(proliferation)과 사멸(apoptosis)에 영향을 미치는 것으로 알려져 있다[ 9, 13, 14, 49]. 운동의 효과는 염증 반응에 긍정적인 변화를 가져오는 것으로 잘 알려져 있으며, 특히 항염증 사이토카인의 분비 및 염증성 사이토카인의 분비 억제를 일으키는 것으로 알려져 있다[ 42]. 따라서 본 연구에서 24주간의 트레드밀 운동의 효과가 염증반응에 대해 긍정적인 변화를 일으켰고 이로 인해 대장암 폴립에서 SPARC의 발현을 유의미하게 증가시킨 것으로 사료된다.
4. 장기간의 트레드밀 운동이 대장암 유발 쥐의 OSM 발현에 미치는 영향
IL-6 family 사이토카인 중 하나인 OSM은 염증 반응에 관여하며 세포의 성장, 항상성 유지, 간 기능, 신경세포 성장과 골 유지를 위해 중요한 역할을 하는 것으로 알려져 있다[ 50]. 또한 OSM는 다양한 조직에서 종양 세포의 증식 억제를 일으키는 것으로 알려져 있는데 [ 15, 51, 52], 암 세포에서는 그 발현이 감소되어 있다[ 53]. 따라서 OSM의 조절은 암의 관리에 있어 중요한 기전으로 여겨지고 있다. 본 연구에서 24주간의 장기간 운동에 의해 비복근에서 OSM의 발현을 살펴본 결과 운동군(EX)에서의 발현이 통제군(CON)에 비해 유의하게 증가하였으나 혈중과 폴립에서는 통계적으로 유의한 차이가 나타나지 않았다. 이러한 결과는 Hojman et al. [ 5]이 수영 운동 직후 해부한 정상 쥐에서 간, 비장 등의 다른 조직에 비해 근육(가자미, 비복근, 경골근 등)에서 특이하게 발현하였다고 보고한 연구결과와 일치한다[ 5]. OSM은 항암 효과를 일으키는 역할로 잘 알려져 있는데, 특히 유방암 세포로 알려진 MCF-7에 있어 의존적(dose-dependent) 효과가 있다고 알려져 있고[ 54], 이는 Transcriptional suppression of synuclein gamma (SNCG) 발현을 통한 종양세포 증식 억제와[ 52, 55], p53, caspase 등의 작용을 통한 종양 세포의 자멸(apoptosis)을 일으키는 것으로 알려져 있다[ 5]. 비록 운동 유발성 마이오카인과 암과의 관계를 규명하고자 하는 대부분의 선행 연구가 배양된 세포를 이용한 실험이거나, 임상 실험들도 일회성 운동 중재이며, 암 종에 따라 관련 변인을 살펴본 연구가 미흡하여 본 연구의 결과와 직접적인 비교를 하는 것에는 제한이 있다. 그러나 본 연구는 일회성 운동에 의한 일시적 발현 증가가 아닌 장기간 운동에 의해 안정 시 수준에서 유의미한 발현을 확인하였다는 것에 의의가 있으며, 또한 OSM의 발현과 관련하여서는 아직까지 유방암, 난소암, 폐암 등의 암종에서만 연구가 되어왔으므로[ 15, 51, 55], 대장암에서 운동에 의해 발현이 증가하였음을 확인하였다는 것에 의미가 있다. 따라서 추후 연구에서는 장기간의 운동 중재와 더불어 다양한 운동 강도와 형태를 적용한 연구들, 암의 증식과 사멸에 관련한 변인들의 변화를 밝히는 연구가 필요할 것으로 생각된다.
결론
본 연구는 6주령의 ICR 수컷 쥐를 대상으로 중강도 트레드밀 운동을 실시하고 이후 암을 유발하여 근육, 혈장, 대장폴립에서 SPARC와 OSM의 발현에 미치는 영향을 조사하였으며 다음과 같은 결론을 얻었다.
장기간 지속된 트레드밀 운동은 암으로 인한 체중 감소를 유의미하게 억제시켰으며, 복부 내장 지방량을 유의하게 감소시켰다. 또한 장기간 지속된 트레드밀 운동은 대장 폴립의 SPARC 발현을 유의하게 증가시켰으며, 근육에서 OSM의 발현을 유의하게 증가시킨 것을 확인할 수 있었다.
본 연구의 결과를 통해 장기간 규칙적인 유산소 운동은 암세포자멸과 증식억제와 관련이 있는 SPARC와 OSM 발현에 영향을 미쳤으며, 이는 암에 대한 운동의 효과를 밝히는 기전 연구에 기초 자료가 될 것으로 사료된다.
Fig. 1.
Fig. 1.The Protocol of the study. AOM, Azoxymethane; DSS, Dextran Sodium Sulfate. 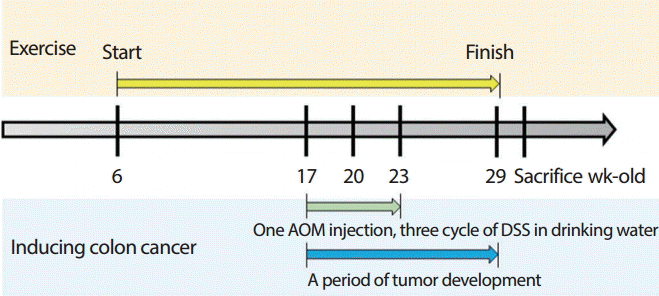
Fig. 2.
Fig. 2.The change of body weight. 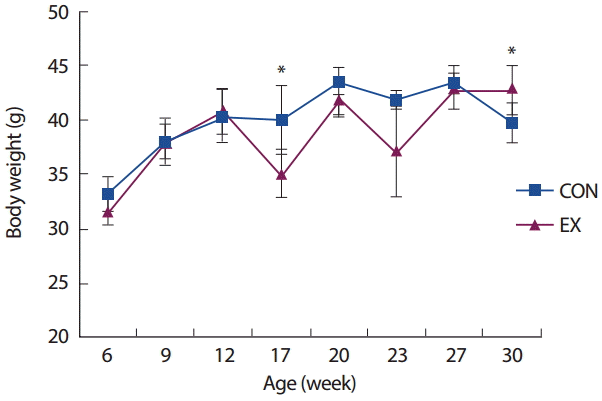
Fig. 3.
Fig. 3.
The expression of SPARC in skeletal muscle (A), blood (B), polyp (C) between control group (CON) and 24-week treadmill exercise group (EX). Values are means±SED.
*: p<.05 significantly different from CON.
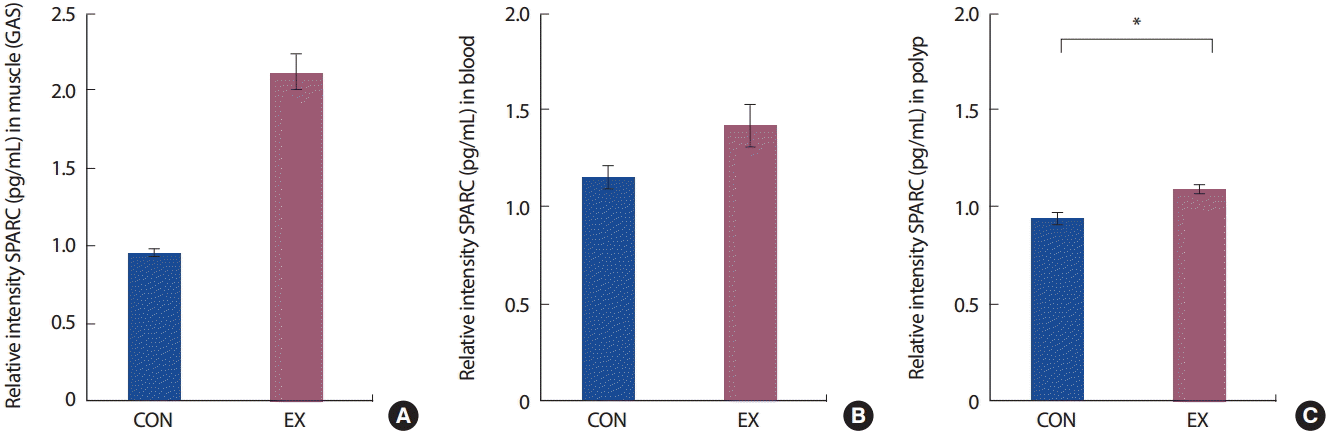
Fig. 4.
Fig. 4.
The expression of Oncostatin M (OSM) in skeletal muscle (A), blood (B), polyp (C) between control group (CON) and 24-week treadmill exercise group (EX). Values are means±SED.
*: p<.01 significantly different from CON.
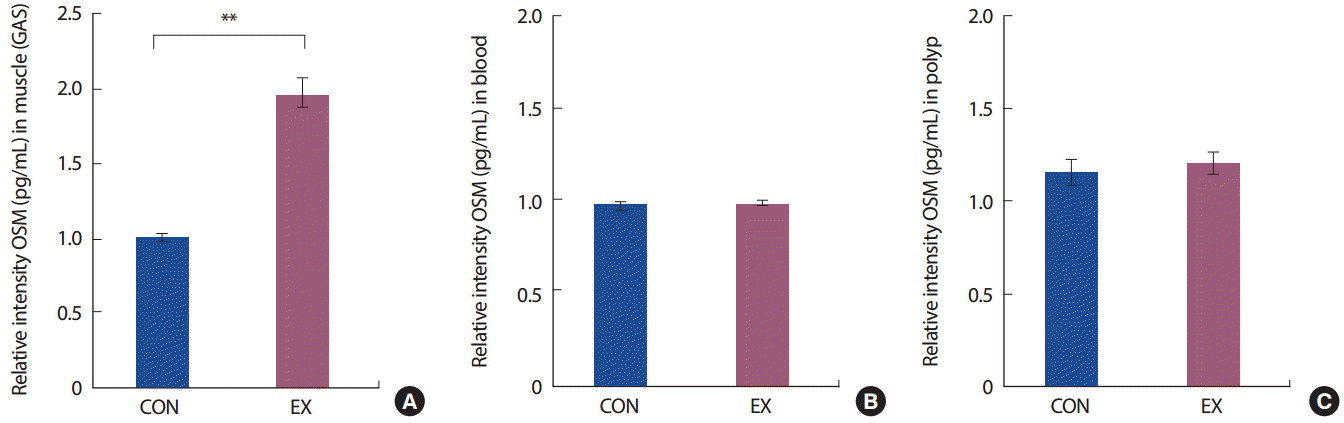
Table 1.
24-week treadmill exercise protocol
|
Age (wk) |
Exercise duration |
Frequency |
Intensity (m/min) |
|
6-7 |
1-2 wk |
30 min/day |
14 |
|
8-13 |
3-8 wk |
5 d/wk |
15-17 |
|
14-29 |
9-24 wk |
|
18 |
Table 2.
The difference of body composition
|
CON |
EX |
p
|
|
Body weight (g) |
39.67 ± 1.83 |
42.72 ± 2.23 |
.026*
|
|
Gastrocnemius (g) |
0.19 ± 0.13 |
0.19 ± 0.01 |
.296 |
|
Quadriceps (g) |
0.22 ± 0.02 |
0.24 ± 0.01 |
.210 |
|
Abdominal fat (g) |
0.70 ± 0.35 |
0.26 ± 0.08 |
.014*
|
REFERENCES
1. Pedersen BK, Febbraio MA. Muscle as an endocrine organ: focus on muscle-derived interleukin-6. Physiological Reviews. 2008;88(4):1379-1406.   2. Pedersen BK, Fischer CP. Physiological roles of muscle-derived interleukin-6 in response to exercise. Current Opinion in Clinical Nutrition and Metabolic Care. 2007;10(3):265-271. 3. Rohde T, MacLean DA, Richter EA, Kiens B, Pedersen BK. Prolonged submaximal eccentric exercise is associated with increased levels of plasma IL-6. The American Journal of Physiology. 1997;273(1 Pt 1):E85-91.
4. Broholm C, Pedersen BK. Leukaemia inhibitory factor--an exercise-induced myokine. Exercise Immunology Review. 2010;16:77-85. 5. Hojman P, Dethlefsen C, Brandt C, Hansen J, Pedersen L, et al. Exercise-induced muscle-derived cytokines inhibit mammary cancer cell growth. American journal of physiology. Endocrinology and Metabolism. 2011;301(3):E504-510.
6. Bornstein P, Sage EH. Matricellular proteins: extracellular modulators of cell function. Current Opinion in Cell Biology. 2002;14(5):608-616. 7. Hasselaar P, Sage EH. SPARC antagonizes the effect of basic fibroblast growth factor on the migration of bovine aortic endothelial cells. Journal of Cellular Biochemistry. 1992;49(3):272-283. 8. Yan Q, Sage EH. SPARC, a matricellular glycoprotein with important biological functions. Journal of Histochemistry and Cytochemistry. 1999;47(12):1495-1506. 9. Tai IT, Tang MJ. SPARC in cancer biology: its role in cancer progression and potential for therapy. Drug Resistance Updates. 2008;11(6):231-246. 10. Bellahcene A, Castronovo V. Increased expression of osteonectin and osteopontin, two bone matrix proteins, in human breast cancer. The American Journal of Pathology. 1995;146(1):95-100.  11. Yang E, Kang HJ, Koh KH, Rhee H, Kim NK, et al. Frequent inactivation of SPARC by promoter hypermethylation in colon cancers. International Journal of Cancer. 2007;121(3):567-575. 12. Park SI, Lee ED, Suh DS, Kim KH. MS. The Clinicopathological Relevance for The Expression of Secreted Protein Acidic and Rich in Cysteine in Epithelial Ovarian Tumors. Obstetrics & Gynecology Science. 2011;54(12):770-777.
13. Aoi W, Naito Y, Takagi T, Tanimura Y, Takanami Y, et al. A novel myokine, secreted protein acidic and rich in cysteine (SPARC), suppresses colon tumorigenesis via regular exercise. Gut. 2012;62(6):882-889. 15. Grant SL, Begley CG. The oncostatin M signalling pathway: reversing the neoplastic phenotype? Molecular Medicine Today. 1999;5(9):406-412. 17. Kim CS. Translated Expression of bone-resorbing cytokines according to aerobic exercise intensities. Journal of Exercise Nutrition & Biochemistry. 2011;15(3):131-139. 18. Kim CW, Kim GD, Kang SH, Park CH, Kim KB, et al. Translated The Effect of 16 Weeks of Resistance Training on the Fatigue Factor, Muscle Soreness, Oxidative Stress, and Myokine in Elite Weightlifters. Korean Society of Life Science. 2012;22(2):184-191. 19. Febbraio MA, Pedersen BK. Muscle-derived interleukin-6: mechanisms for activation and possible biological roles. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2002;16(11):1335-1347. 20. Tamura Y, Watanabe K, Kantani T, Hayashi J, Ishida N, et al. Upregulation of circulating IL-15 by treadmill running in healthy individuals: is IL-15 an endocrine mediator of the beneficial effects of endurance exercise? Endocrine Journal. 2011;58(3):211-215. 21. Neufert C, Becker C, Neurath MF. An inducible mouse model of colon carcinogenesis for the analysis of sporadic and inflammation-driven tumor progression. Nature Protocols. 2007;2(8):1998-2004. 22. Bedford TG, Tipton CM, Wilson NC, Oppliger RA, Gisolfi CV. Maximum oxygen consumption of rats and its changes with various experimental procedures. Journal of Applied Physiology. 1979;47(6):1278-1283. 23. Solymoss BC, Bourassa MG, Lesperance J, Levesque S, Marcil M, et al. Incidence and clinical characteristics of the metabolic syndrome in patients with coronary artery disease. Coronary Artery Disease. 2003;14(3):207-212. 24. Wheelwright S, Darlington AS, Hopkinson JB, Fitzsimmons D, White A, et al. A systematic review of health-related quality of life instruments in patients with cancer cachexia. Supportive Care in Cancer. 2013;21(9):2625-2636. 25. Bachmann J, Heiligensetzer M, Krakowski-Roosen H, Buchler MW, Friess H, et al. Cachexia worsens prognosis in patients with resectable pancreatic cancer. Journal of Gastrointestinal Surgery. 2008;12(7):1193-1201. 26. Meriggi F. Cancer Cachexia: one step ahead. Reviews on Recent Clinical Trials. 2015;10(3):246-250. 27. Giordano A, Calvani M, Petillo O, Carteni M, Melone MR, et al. Skeletal muscle metabolism in physiology and in cancer disease. Journal of Cellular Biochemistry. 2003;90(1):170-186. 28. Woods JA. Exercise and resistance to neoplasia. Canadian Journal of Physiology and Pharmacology. 1998;76(5):581-588. 29. L IS. Translated The Approach of Antioxidant, Endocrine, & Immune System on Exercise & Aging. Exercise Science. 2002;11(1):35-51.
30. Ardies CM. Exercise, cachexia, and cancer therapy: a molecular rationale. Nutrition and Cancer. 2002;42(2):143-157. 32. Zhou X, Wang JL, Lu J, Song Y, Kwak KS, et al. Reversal of cancer cachexia and muscle wasting by ActRIIB antagonism leads to prolonged survival. Cell. 2010;142(4):531-543. 33. Daneryd PL, Hafstrom LR, Karlberg IH. Effects of spontaneous physical exercise on experimental cancer anorexia and cachexia. European Journal of Cancer and Clinical Oncology. 1990;26(10):1083-1088. 34. Baltgalvis KA, Berger FG, Pena MM, Davis JM, Carson JA. Effect of exercise on biological pathways in ApcMin/+ mouse intestinal polyps. Journal of Applied Physiology. 2008;104(4):1137-1143. 35. Colbert LH, Mai V, Perkins SN, Berrigan D, Lavigne JA, et al. Exercise and intestinal polyp development in APCMin mice. Medicine and Science in Sports and Exercise. 2003;35(10):1662-1669. 37. Lee IM, Paffenbarger RS Jr. Quetelet’s index and risk of colon cancer in college alumni. Journal of the National Cancer Institute. 1992;84(17):1326-1331. 38. Murphy TK, Calle EE, Rodriguez C, Kahn HS, Thun MJ. Body mass index and colon cancer mortality in a large prospective study. American Journal of Epidemiology. 2000;152(9):847-854. 39. Kim MC, Kim CS. TH. Translated The Effect of Physical Activity on Colorectal Polyps. Korean Journal of Family Medicine. 2005;26(7):17-22.
40. Park HS, Park JY, Yu R. Relationship of obesity and visceral adiposity with serum concentrations of CRP, TNF-alpha and IL-6. Diabetes Research and Clinical Practice. 2005;69(1):29-35. 41. Lee HL, Son BK, Lee OY, Jeon YC, Han DS, et al. Translated Abdominal Obesity, Insulin Resistance, and the Risk of Colonic Adenoma. The Korean Journal of Gastroenterology. 2007;49(3):147-151. 42. Kondo T, Kobayashi I, Murakami M. Effect of exercise on circulating adipokine levels in obese young women. Endocrine Journal. 2006;53(2):189-195. 43. Kim KJ. The associations of obesity and exercise participation with body composition. blood revels of IL-6 and TNF-a in adult women. Exercise Science. 2008;17(2):119-128. 44. Winters-Stone KM, Dieckmann N, Maddalozzo GF, Bennett JA, Ryan CW, et al. Resistance Exercise Reduces Body Fat and Insulin During Androgen-Deprivation Therapy for Prostate Cancer. Oncology Nursing Forum. 2015;42(4):348-356. 46. Kos K, Wilding JP. SPARC: a key player in the pathologies associated with obesity and diabetes. Nature reviews. Endocrinology. 2010;6(4):225-235. 47. Yang EG. Identification and clinical signification of SPARC inactivation through promoter hypermethylation in human colorectal cancers [dissertation]. 2006;Yonsei University, Seoul.
48. Podhajcer OL, Benedetti LG, Girotti MR, Prada F, Salvatierra E, et al. The role of the matricellular protein SPARC in the dynamic interaction between the tumor and the host. Cancer Metastasis Reviews. 2008;27(4):691-705. 49. Chen J, Wang M, Xi B, Xue J, He D, et al. SPARC is a key regulator of proliferation, apoptosis and invasion in human ovarian cancer. PloS One. 2012;7(8):e42413. 50. Heinrich PC, Behrmann I, Haan S, Hermanns HM, Muller-Newen G, et al. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. The Biochemical Journal. 2003;374(Pt 1):1-20. 51. Hutt JA, DeWille JW. Oncostatin M induces growth arrest of mammary epithelium via a CCAAT/enhancer-binding protein delta-dependent pathway. Molecular Cancer Therapeutics. 2002;1(8):601-610. 52. Liu J, Spence MJ, Zhang YL, Jiang Y, Liu YE, et al. Transcriptional suppression of synuclein gamma (SNCG) expression in human breast cancer cells by the growth inhibitory cytokine oncostatin M. Breast Cancer Research and Treatment. 2000;62(2):99-107. 53. Garcia-Tunon I, Ricote M, Ruiz A, Fraile B, Paniagua R, et al. OSM, LIF, its receptors, and its relationship with the malignance in human breast carcinoma (in situ and in infiltrative). Cancer Investigation. 2008;26(3):222-229. 54. Douglas AM, Grant SL, Goss GA, Clouston DR, Sutherland RL, et al. Oncostatin M induces the differentiation of breast cancer cells. International Journal of Cancer. 1998;75(1):64-73. 55. Liu J, Spence MJ, Wallace PM, Forcier K, Hellstrom I, et al. Oncostatin M-specific receptor mediates inhibition of breast cancer cell growth and down-regulation of the c-myc proto-oncogene. Cell Growth & Differentiation: the Molecular Biology Journal of the American Association for Cancer Research. 1997;8(6):667-676.
|
|





