서 론
다양한 운동 강도와 형태 중 간헐적 운동 및 일회성의 중강도 유산소 운동은 대표적인 운동 형태이다[1]. 이러한 일회성 운동은 세포내 에너지 수준의 변화와 근육의 수축 작용 등을 일으키게 되는데 이러한 변화가 자극원이 되어 호르몬 및 세포내 신호 전달을 통해 인체의 분자 생리적인 반응을 유도한다[2,3]. 운동 자극에 의한 인체의 다양한 반응 중에서 에너지 대사와 관련한 반응은 현대 사회에 만연한 비만이나 이상지질과 같은 대사 질환과 밀접한 관련이 있어 그 의미가 중요하다[4-6].
지금까지 운동자극과 관련하여 다양한 연구들이 진행되어 왔다. 운동이 Malonyl CoA, 근 소포체와 관련된 대사경로의 변화를 유도하고, 근세포나 간세포 내의 지질 수준의 변화에도 영향을 미친다는 것이 발표되었다[7,8]. 또한 사람을 대상으로 한 연구에서는 근세포내 지질축적 정도가 운동 트레이닝에 의해 변화된다는 결과가 있었다[9]. 최근에는 대사와 염증관련 적응의 측면에서 서로 다른 강도의 장단기 운동이 혈중 IL-5, IL-10, TNF-alpha, IL-6/IL-10, NEFA, 글루코스 등의 수준에 긍정적인 영향을 끼친다는 연구와 같이[10] 운동 자극에 대한 변화를 다양한 관점에서 관찰하는 연구가 많이 시행되고 있다. 그러나 다양한 운동 조건에 따른 대규모의 유전자 발현 분석은 미흡한 실정이며 운동자극의 메커니즘적 이해를 위해서 계속적으로 연구되어야 할 필요성이 있다.
이와 같은 운동 자극과 신체적인 적응을 매개함에 있어 근육은 중요한 요소이다. 근육이 우리 몸의 40% 이상을 차지하는 조직이고[11] 핵수용체와 연관된 기전으로 직접적인 에너지 대사 조절 및 마이오카인을 통해 대사를 조절하는 중요한 물질대사 중재 기관이라는 것을 고려할 때[12], 근육에서의 유전자 발현변화는 생리적 항상성에 매우 중요하다고 할 수 있다. 그리고 이러한 근육조직에서의 유전자 발현변화는 결국 에너지 대사에 중요한 이동통로 단백질이나 효소들의 활성을 조절하는 분자적 기전[13,14]의 시초가 된다.
한편, 지금까지 많은 연구가 일회성 운동의 유전자 발현과 관련하여 이루어져 왔고[15-17] 건강한 마우스에서 2-15일의 운동이 연골조직에 미치는 영향을 microarray 방식으로 분석한 연구도 있다[18]. 그러나 정상 상태의 마우스에게 일상적으로 일어나는 짧은 일회성의 최대하 운동을 시행하고 이를 microarray 기술을 사용함으로써 마우스의 비복근 근육의 전체적인 유전자 발현을 에너지 대사의 관점에서 관찰한 사례는 상대적으로 많지 않은 실정이다. 전체적인 유전자 발현 관찰에 있어서 본 연구에서 이용한 microarray 분석기법은 운동 자극에 대한 유전자의 전체적인 발현을 대규모로 살펴볼 수 있고, 관련된 특이적인 유전자의 발현을 심층적으로 분석할 수 있는 토대가 된다[19]. 아울러 일회성 운동은 운동 자극의 출발점이 되는 동시에 생리적 운동 적응의 시발점이 되는 만큼[20] 다양한 방법으로 연구되어져야 한다.
따라서 본 연구는 일상적으로 행해지는 형태의 일회성 유산소성 운동 후 비복근 근육에서의 전체적인 유전자 발현을 microarray 기법을 이용하여 분석하였다. 또한 도출된 차별발현 유전자(DEGs)를 에너지 대사 및 운동적응의 관점에서 바라보고 기존의 알려진 연구들과 비교하여 확인하는 과정을 진행하였다. 그리고 Atf3 유전자가 운동자극에 의해 발현 수준이 변화되고 운동적응에 역할을 할 수 있다는 연구와 같이[21], 본 연구의 microarray 분석은 근육에서 에너지 대사와 관련된 유전자의 초기 발현 수준에 일회성 운동이 미치는 영향을 구체적으로 검토하였다.
또한 최근에는 운동관련 분야에서도 생물정보학을 이용한 빅데이터 분석 관련 연구가 진행되고 있는데[22] 본 연구에서도 microarray 분석 결과와 함께 생물정보학 도구인 DAVID의 정보를 비교 및 검증해 보면서 발현의 변화가 있는 유전자들을 군집화하고 Gene ontology와 KEGG pathway를 바탕으로 한 분석을 진행하였다. 이러한 분석들을 바탕으로 1시간의 지정된 프로토콜의 일회성 운동이 근육에서 에너지 대사의 적응과 관련한 유전자의 초기 발현 수준에 미치는 영향을 살펴보고자 한다.
연구 방법
1. 실험동물
본 연구에 사용된 실험동물(Strain: C57BL/6N) 8주령 수컷 쥐 10마리는 중앙실험동물㈜로부터 구입하여 서울대학교 수의학과의 동물실험실의 정책 아래 관리되었다. 멸균된 물과 사료는 규칙적으로 공급하였으며 접근은 자유로웠다. 모든 마우스는 섭씨 22°C, 40-50%의 습도, 밤낮 주기(12시간 점등/12시간 소등)에서 사육되었다. 동물실험의 승인은 서울대학교 IACUC 위원회의 승인(과제번호: SNU-170518-9)을 받았으며 마우스들은 전용케이지에 각각 5마리씩 무작위적으로 배정되어 대조군과 운동 그룹으로 나누어 사육하였다.
2. 조직적출
마우스는 Isoflurane으로 흡입마취를 한 후 심장 채혈을 실시하였다. 마취부터 조직적출까지의 과정은 5분 이내로 실시하였으며 일회성 운동의 효과를 보기 위하여 운동 측정 종료 후 2시간 이내에 적출을 실시하였다. 적출 근육은 무게(Wet weight)를 측정하고 즉시 Cryotube에 넣어 액체질소에 급 냉각하여 -80°C에 보관하였다.
3. 운동 처치
운동군에 속한 마우스는 Exer 3/6 Columbus instruments를 이용하여 5일간의 트레드밀 적응 운동을 실시하고 매일 몸무게와 식이 섭취량을 기록하였다. 일회성 운동의 시간과 강도는 마우스를 대상으로 한 선행연구를 바탕으로 변형하여 실시하였다[23-26]. 초기 0-10분은 10 m/min으로 설정한 후, 10-20분까지 2분마다 2 m/min씩 점진적으로 속도를 증가시켜 18 m/min에 도달하도록 하였다. 20-25분까지 18 m/min을 유지한 후 25-60분까지 16 m/min으로 마무리하였다. 경사도는 0%였다. 운동 도중 마우스는 부드럽게 손으로 두드려 측정시간 동안 계속 하여 달리도록 하였다.
4. Microarray 분석과 자료처리
RNA의 purity와 integrity는 ND-1000 Spectrophotometer (Nano-Drop, Wilmington, USA), Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, USA)를 통해 확인되었다. 진행된 샘플은 Acute exercise (AC)1, AC3, AC4와 Control (Con)1, Con2, Con3, Con4로 전체 7개의 샘플을 두 그룹으로 나누어 비교하였다. AC2 샘플은 RIN 값과 rRNA ratio값이 각각 5.2, 0.6으로써 Quality Check 기준을 통과하지 못하여 분석에서 제외하였다. Array 분석은 Affymetrix사의 Whole transcript Expression Arrays Methods를 사용하였다. The Affymetrix Whole transcript Expression array 과정은 제조사(Gene Chip Whole Transcript PLUS reagent Kit)의 프로토콜을 따라 시행되었다. cDNA는 Gene Chip Whole Transcript (WT) Amplification kit를 사용하여 제조사의 방법에 따라 합성되었으며, 혼성화된 array는 GCS3000 Scanner (Affymetrix)를 사용하여 스캔하였다. 신호 값들은 AffymetrixⓇ Gene ChipTM Command Console software로 계산하였다. 결과 데이터는 gene level RMA 분석을 하였고 차별발현 유전자 분석을 하였다.
발현 데이터의 통계적 유의성은 그룹 간 차이가 없을 것이라는 귀무가설을 바탕으로 local-pooled-error (LPE) test와 fold change 기준으로 진행하였다(Cut-off 기준: 비교조합에서 fc >_1.5인 probes). 차별발현 유전자 집합은 Hierarchical cluster analysis를 진행하였고, 유의한 발현의 정도를 보인 유전자들 대상으로는 Gene enrichment와 Functional annotation analysis를 각각 Gene ontology와 KEGG pathway 분석을 바탕으로 진행하였다. 유의한 발현을 보인 유전자들의 모든 데이터 분석과 시각화는 R software를 사용하여 진행하였다.
연구 결과
1. Gene Ontology와 KEGG pathway를 바탕으로 한 Microarray data의 gene-enrichment와 functional annotation 분석
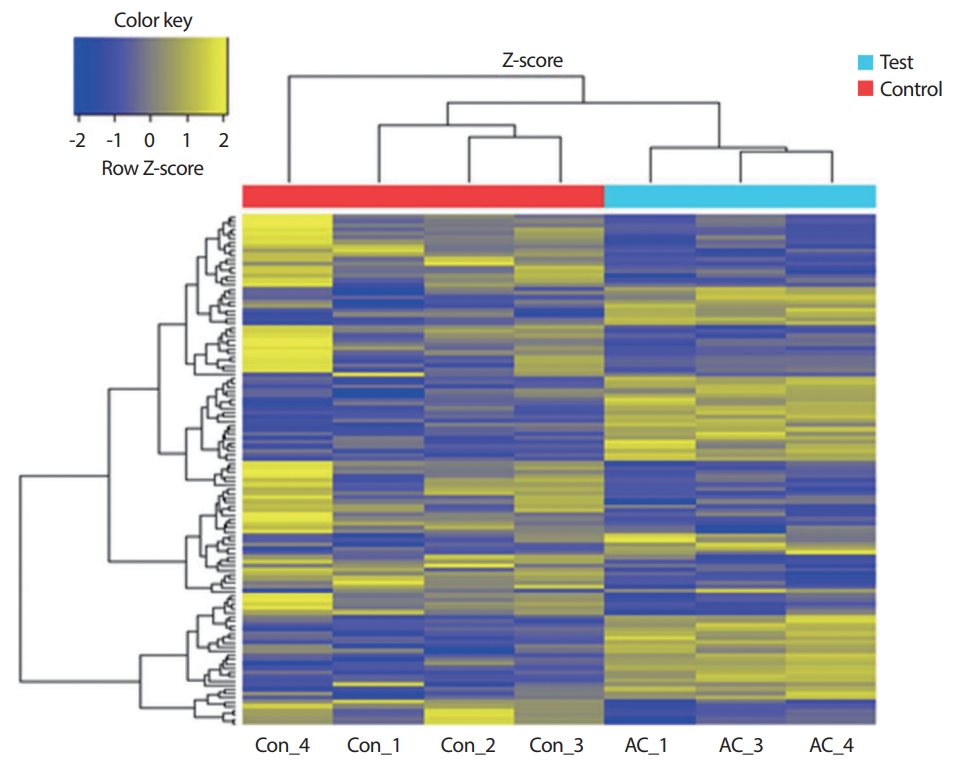
유의한 발현량의 차이를 가지는 차별발현 유전자를 대상으로 한 microarray 실험 결과, 대조군 대비 운동그룹에서 상향 조절(up-regulation)된 유전자는 55개, 하향 조절(down-regulation)된 유전자는 65개였다. 그중 상위 10개의 상향, 하향 조절된 유전자는 Table 1과 같다. Hierarchical 분석을 통한 그룹 간 전체 microarray chip 유전자의 발현 패턴의 경우 대조군과 운동군 간에는 뚜렷한 발현 패턴의 차이를 보이며 대조군 그룹, 운동군 그룹 내에서는 유사성이 관찰되었다(Fig. 1).
1) Gene ontology (GO) 데이터베이스를 기반으로 한 enrichment 분석(Biological Process category)
일회성 운동 처치 후 유의한 발현 수준의 차이를 보이는 차별발현 유전자들을 대상으로 한 본 연구의 ontology 분석 결과, Biological Process 범주에서는 cellular process, biological regulation, metabolic process, regulation of biological process, regulation of cellular process, response to stimulus, cellular metabolic process 등이 enrichment 분석에서 유의수준을 기준으로 나열한 상위 10개의 유의한(p<.001) GO용어였으며 각각 60개 이상의 차별발현 유전자를 포함하였고 이 중 가장 많은 차별발현 유전자가 연결된 용어는 cellular process였다(Fig. 2). 이는 일회성 운동 자극이 cellular process와 밀접한 관련을 가지고 있다는 것을 보여준다.
2) Gene ontology 데이터베이스를 기반으로 한 enrichment 분석(Cellular Component category)
Cellular component와 관련된 enrichment 분석에서는 cell part의 intracellular, intracellular part, cytoplasm/cytoplasmic part, membranebounded organelle, intracellular membrane-bounded organelle 등의 GO 용어가 유의하였으며(p <.001) 각각 40개 이상의 차별발현 유전자가 연결되었고 가장 많은 차별발현 유전자가 연결된 용어는 cell part였다(Fig. 3).
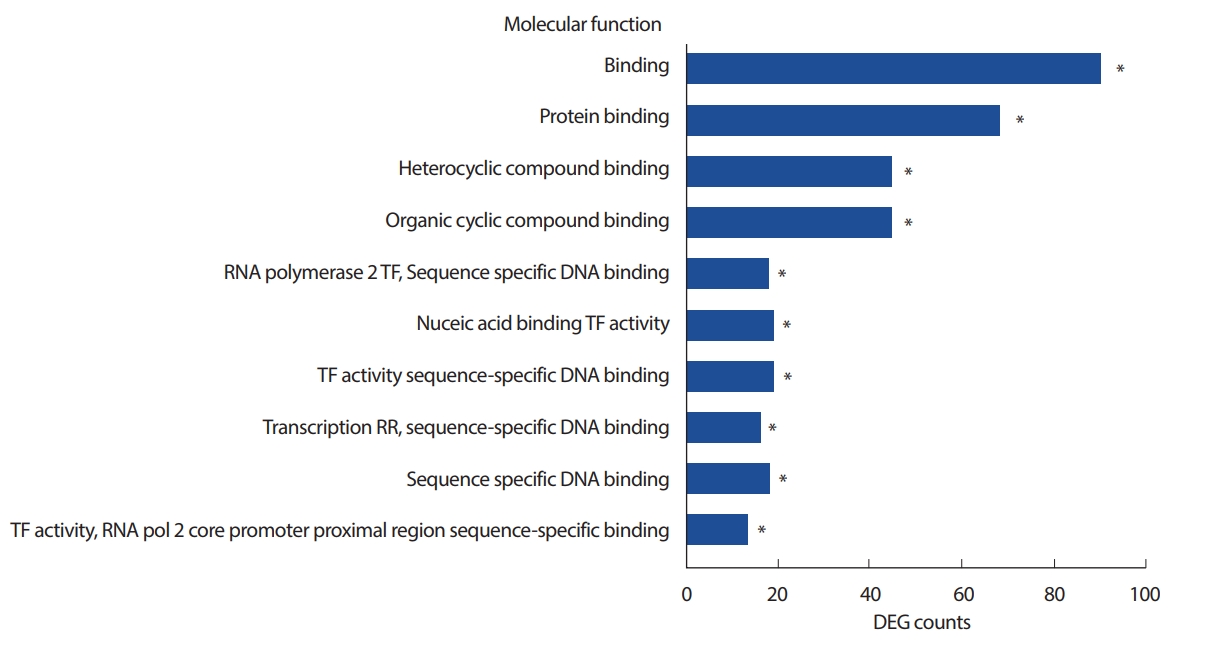
3) Gene ontology 데이터베이스를 기반으로 한 enrichment 분석(Molecular process category)
Molecular process와 관련된 enrichment 분석에서는 주로 binding과 전사활성, 전사조설 등과 관련이 있는 binding, protein binding 용어 등에 각각 40개 이상의 차별발현 유전자가 연결되었고 해당 GO 용어들은 통계적으로 유의하였다(p<.001)(Fig. 4).
4) 에너지 대사 관점의 Microarray data KEGG pathway 결과 분석
본 연구의 차별발현 유전자를 대상으로 KEGG pathway database에 기반한 KEGG Enrichment 분석을 실시하였고 그중 유의미한 에너지 대사관련 pathway 용어들을 선별하였다(Table 2). Global and overview maps 측면에서는 metabolic pathways가 유의하게(p <.01) 나타났다. Biosynthesis of other secondary metabolites 범주에서는 neomycin, kanamycin and gentamicin biosynthesis가 유의하게(p <.05) 나타났으며 Genetic information processing에서는 folding, sorting and degradation 범주의 protein processing in endoplasmic reticulum 관련 pathway가 유의하였다(p<.05). 구체적으로 단백질 분해와 세포질의 ubiquitin ligase complex에서 역할을 하는 Heat shock protein 계열인 hspa1a, b와 hspa2, dnajb1 등이 대조군 대비 1.5배 이상 발현정도가 증가하였다. Environmental information processing에서는 몇몇의 signal transduction 관련 pathway가 유의하였는데, MAPK signaling pathway의 경우 그 유의도(p <.01)가 매우 높았다. 이러한 높은 유의도는 세포의 증식과 분화와 관련이 있는 c-fos유전자(2.83배 증가)와 c-jun유전자(1.51배 증가)에서 구체적으로 관찰되었다. 이밖에 Jak-STAT, TNF, HIF-1 signaling pathway (p <.05)와 함께 운동 자극에 의해 유도되는 AMPK signaling에서는 AMPK의 상위 수준에서 기능을 할 수 있는 렙틴(-1.54배 감소)과 하위 수준에 있는 SCD3 유전자(-1.63배 감소)에서 유의하게(p<.01) 나타났다.
Cellular Process의 transport and catabolism 범주에서는 phagosome과 cell growth and death 부분과 같은 apoptosis 관련 pathway가 유의하였다(p<.05). Organismal system에서는 immune system과 endocrine system에서 몇몇의 pathway enrichment가 나타났는데, Toll-like receptor signaling pathway (p <.05), NOD-like receptor signaling pathway (p<.01), Th17cell differentiation (p<.05)이 유의하였고 antigen processing and presentation과 IL-17signaling pathway의 경우는 매우 유의하였다(p<.001).
에너지 대사와 관련이 있는 endocrine system의 경우, Insulin signaling pathway (p<.05), adipocytokine signaling pathway (p <.01), PPAR signaling pathway (p<.01)가 유의하였다. 특히 Estrogen signaling pathway는 그 유의도가 높았다(p <.001). 한편 에너지 대사와 관련된 또 다른 pathway인 carbohydrate metabolism 범주에서는 Glycolysis/gluconeogenesis (p<.01)가 유의하였는데 이 pathway에서 역할을 하는 HK1, HK2, Adh1 등의 유전자들이 대조군 대비 유의한 발현 수준의 변화가 나타났다. 구체적으로 해당과정의 시작과 핵산의 당 신생, 과당과 만노오스와 같은 탄수화물 대사와 관련 있는 hexokinase1 (HK1)은 감소(-1.5배 감소)하였지만 다른 Fructose나 mannose, galactose, starch, sucrose, amino sugar, nucleotide 관련 metabolism은 KEGG pathway 데이터 기반 enrichment 분석에서 유의하지 않게 나타났다.
2. The Database for Annotation, Visualization and Integrated Discovery (DAVID)를 이용한 본 연구의 microarray 분석의 검증 및 비교
상향조절된 차별발현 유전자 56개(1.5 fold 기준)를 대상으로 하여 DAVID 분석을 진행하였다.
1) Gene ontology (Biological Process) 분석
DAVID의 Biological process 범주와 관련하여서 가장 많은 관련 차별발현 유전자들이 속해 있는 GO용어는 순서대로 regulation of transcription, DNA-templated (19개), transcription, DNA-templated (17개), negative/positive regulation of transcription from RNA polymerase 2 promoter (13개, 10개), regulation of transcription from RNA polymerase 2 promoter (9개), negative regulation of apoptotic process (9개), positive regulation of transcription, DNA-templated (8개), transcription from RNA polymerase 2 promoter (7개) 등으로서 전사와 전사조절과 관련된 용어들이 있었다. 본 연구의 enrichment 분석과 DAVID분석 결과와 비교하여 볼 때 공통적으로 세포 수준에서의 생리학적인 조절, 특히 대사과정의 전사조절과 관련된 유전자의 발현이 대조군 대비 유의하였다.
2) Gene ontology (Cellular Component) 분석
DAVID의 Cellular Component 범주 관련 GO 용어 중 가장 많은 차별발현 유전자들이 속해 있는 용어는 nucleus (26개), cytoplasm (25개)이었다. 이는 본 연구의 enrichment 분석에서 세포안의 핵이나 세포질, 세포막/핵막과 붙어있는 기관들과 관련된 유전자들이 많다는 것과 유사하다.
논 의
본 연구에서는 일회성 운동 자극 후 마우스 비복근의 전체적인 유전자 발현변화를 관찰하였다. 도출된 차별발현 유전자는 분석을 통해 기존의 생물학적 기반 GO 용어기준으로 클러스터링(clustering)하였다. 아울러 본 연구에서는 microarray 검증 및 분석 도구로서 활용한 DAVID 분석을 통해 전통적인 통계기법을 사용한 분석보다 많은 수의 차별발현 유전자를 대상으로 하여 functional annotation, classification 분석을 할 수 있었다[27]. 이러한 분석 결과는 개별 유전자의 발현 변화만으로는 발견할 수 없는 의미를 전체적인 관점에서 해석함으로써 운동자극으로 나타나는 생리적인 변화를 해석하는 데 도움을 줄 수 있다.
본 연구의 enrichment 분석에서는 조절, 자극, 대사관련 용어들이 상위 10개의 Biological Process GO용어에 유의미하게 나타났다(Fig. 2). 유의미하다는 것은 마우스의 전체 유전체에서 해당 GO용어에 연결/연결되지 않은 유전체의 비율과 본 연구의 차별발현 유전자내에서 연결/연결되지 않은 유전체의 비율의 통계적 차이를 의미한다. 또한 Molecular function 범주에서는 binding, protein binding 용어들이 각각 80개, 60개 이상의 차별발현 유전자들을 포함함으로써 유의미하게 상위 1, 2위로 관찰되는 것으로 보아(Fig. 4), 일회성 운동 자극을 통하여 도출된 차별발현 유전자는 물리적으로 분자들 사이에서 접촉하여 기능을 하는 과정과 관련한 역할을 할 수 있다고 사료된다. 이들 차별발현 유전자는 세포 핵산의 전사조절이나 효소 단백질의 조절과 같은 분자적 기전에 영향을 끼칠 수 있는 가능성이 있다.
KEGG pathway 분석 측면에서는, metabolic signaling pathway가 대조군 대비 변화양상이 통계적으로 유의하였고(Table 2), 특히 MAPK pathway가 유의미한 것은 DAVID의 결과와 microarray KEGG enrichment 분석이 일치하였다. KEGG pathway에서 MAPK pathway는 receptor mediated signaling으로서 많은 cellular function 특히 cell proliferation, differentiation and migration과 연관이 있었다. Akimoto et al. [28]에 따르면 C2C12 세포에 mitogen인 p38이 과 발현되었을 때 MAPK 신호전달이 활성화되었고 미토콘드리아의 에너지 대사와 관련이 있는 PGC-1 alpha의 프로모터 활성이 높아졌다고 하였다. 또한 MAPK pathway는 항-세포사멸, 염증반응, 세포주기와 관련이 있었고 JNK and p38 MAP kinase pathway, ERK5 pathway 등과 연결되어 있었다. 이러한 pathway들은 하위 분자들을 통해 각각 p53 signaling pathway, Wnt signaling pathway 등과도 관련이 있고 이들 pathway가 운동에 의해 조절될 수 있다는 연구가 밝혀지고 있는 만큼[29,30] 일회성 운동 자극이 어떠한 구체적인 경로를 통해 에너지 대사에서 역할을 할 수 있는지 추가적인 연구가 필요하다.
한편 본 연구의 microarray 결과로부터 새롭게 도출된 차별발현 유전자 집합 중 상위 10개, 하위 10개의 발현 차이를 보인 유전자들은(Table 1), 선행연구를 토대로 볼 때, 물질대사에서 그 역할을 할 수 있고 나아가 운동 자극과 생리 반응을 매개할 수 있는 가능성이 있다. 먼저 Nr4a3의 경우 본 연구에서 대조군 대비 가장 큰 변화가 관찰되었다. (약 5.85배 증가). Pearen et al. [31]에 따르면, Nr4a 하위 그룹은 핵 수용체로 기능을 하는데 근육에서 beta adrenergic 신호전달을 통하여 글루코스나 인슐린 그리고 지질/에너지 대사에서 역할을 한다고 알려져 있고 최근 연구인 Pearen et al. [32]에서는 증가된 Nr4a3는 운동 적응에 기여를 할 수 있는 다른 유전자의 발현에 영향을 끼칠 수 있다고 하였다. 또한 Liu et al. [33]의 연구에 따르면, L6 근세포의 6-Mercaptopurine의 처리는 인슐린의 존재에 상관없이 NR4A3에 의존적으로 GLUT4의 세포 표면으로의 이동을 통한 글루코스 이동을 향상시킨다고 하였다. 글루코스가 초기 운동반응에서 근육의 중요한 연료로 사용된다는 점을 고려할 때 운동자극으로 유도되는 NR4A3의 초기발현의 증가는 특히 중요하다고 판단된다. 그뿐만 아니라, siRNA로 NOR-1 (NR4A3)을 억제할 때 지방산 산화와 무산소성 대사에서 글루코스/피루빈산의 이용이 악화되고 활성화된 NOR-1의 발현은 지근, 속근 형태의 두 가지 근육에서 산화적 물질대사를 조절한다고도 알려져 있다[34]. 이러한 결과들은 NOR-1이 물질 대사 초기 조절에 깊게 연관될 수 있는 가능성을 시사한다. 두 번째로 가장 많이 변한 Nr4a2 (4.96배 증가)의 경우 Nr2a3와 비슷한 기능을 하는 하위그룹이라는 것이 알려져 있다. 두 유전자 모두 대조군과 비교하여 급격하게 증가한 것을 볼 때, Nr4a 하위그룹은 일회성의 유산소 운동 자극에 가장 민감하게 반응한다는 것을 알 수 있고 운동자극과 생리적 물질대사 반응의 중계자 역할을 할 수 있을 것으로 사료된다.
Btg2 (3.5배 증가)는 본 연구에서 일회성 운동 자극에 의해 대조군대비 매우 민감하게 변하였다. 지방선구세포의 증식과 근육내의 지방조직 성숙에 Btg2가 부정적으로 영향을 끼칠 수 있고[35] 배측 최장근에서는 증가된 Btg2의 발현이 지방축적의 감소를 일으킨다는 연구를 볼 때[36], Btg2와 생성물은 신체 지방의 절대적인 질량 감소에 역할을 할 수 있는 가능성이 있다. 다음으로 증가폭이 큰 Atf3 (3.18배 증가)의 경우 증가된 Atf3의 수준이 아디포넥틴 신호전달의 교란으로 인한 AdipoR1의 감소와 연관이 있다는 연구 결과[37]가 있고 이 유전자는 일회성 운동에 의해 영향을 받는다고 알려졌지만[38,39] 아직 정확한 타겟이 되는 유전자에 대한 연구는 불충분한 상태이다.
이밖에 Otud1, Sik1, Mss51, Thbs1, Irs2, Bcl6 등과 같은 유전자들이 본 연구에서 도출된 차별발현 유전자 중 상위 10개의 상향조절된 유전자들에 포함되어 있었다. 먼저, Otud1은 protease관련 유전자로서 smad7에 작용하여 유방암 전이를 억제할 수 있다는 연구[40]가 있으나 운동과 에너지 대사 관련 역할은 아직 연구가 미비하다. 아울러 인슐린 저항성 마우스 모델에서는 CREB/CRTC2의 활성이 관찰되는데[41] Sik1은 CREB/CRTC2를 억제함으로써 인슐린 저항성을 개선시킨다고 알려져 있다[42]. Mss51은 증식되는 근육과 당분해근육에서 많이 발현되고 대사활성에 부정적인 연관이 있다고 밝혀져 있으며[43] Thbs1을 knockout시킨 마우스는 인슐린 민감도가 높아짐과 동시에 에너지 소비가 증진되었다는 연구 결과가 있다[44]. 그러나 본 연구에서는 일회성의 운동으로 근육에서 Thbs1이 증가된 것으로 나타났고 일반적으로 운동이 인슐린 민감도를 개선한다는 점을 고려할 때 근육에서의 Thbs1의 역할은 추후 연구되어야 한다고 사료된다. 아울러 Irs2의 경우, 골격근에서 베타 산화에 영향을 끼칠 수 있음이 밝혀져 있다[45]. 마지막으로 Bcl6는 최근 근육의 물질대사관련 신호전달에 역할을 한다는 것이 보고되었다[46]. 또한 인간 대상 연구에서는 Bcl6가 최대산소 섭취량과 부적 관계가 있다는 것이 알려져 있다[47]. 본 연구에서는 Bcl6의 발현 수준이 증가되었는데 선행연구와는 상반되는 결과이고 특히 근세포에서 Bcl6의 에너지 대사 측면에서의 역할과 관련한 심화된 연구가 필요하다고 생각된다. 종합하여 볼 때, 이들 유전자들은 에너지 대사와 관련한 역할을 할 수 있을 것으로 유추할 수 있으나 특히 운동과 관련하여 근육에서 구체적으로 어떠한 기능을 할 수 있는지는 연구가 더욱 필요하다.
한편 에너지 대사와 관련하여 대조군 대비 하향 조절된 대표적 유전자에는 Ube2l6와 Scd3가 있었다. Ube2l6와 관련한 연구로서 Ube216의 발현이 감소된 지방세포에서는 지방생성이 억제되어 TG의 accumulation이 줄어드는 것과 관계가 있다는 결과가 발표되었다[48]. 본 연구에서는 Ube216이 대조군대비 약 1.79배 감소되었는데 이는 운동 자극이 UBE2L6의 발현을 감소시켜 지방생성의 초기 단계를 억제하고 나아가 Ube216관련 신호전달에 방해를 일으켜 TG의 축적 또한 억제할 수 있는 가능성을 나타낸다. 다만 본 연구의 모델은 비만, 당뇨 모델이 아닌 정상 상태의 마우스이고 단일 지방세포에서 효과를 본 것이 아니므로 정상적인 생리 상태에서 일회성 운동 자극에 대한 UBEL6의 역할과 그 타겟이 되는 다른 단백질은 추후 연구할 필요성이 있다.
다음으로 Scd3의 경우 대조군 대비 그 발현량이 낮았다(-1.63배 감소). Scd3는 마우스의 경우 Scd (1-4)의 isoform이 있으며 Scd3의 isoform인 Scd1의 경우 짧은 지방산 사슬을 단일 불포화 지방산으로 바꾸는 데 관여한다[49]. Scd3의 에너지 대사관련 연구로서 Stamatikos et al. [50]에 따르면 근육에서 증가된 Scd1의 발현은 saturated fatty acid (SFA)와 SFA로부터 유래된 생성물로 인해 야기된 지질 독성을 줄임과 동시에 심폐적 기능적으로 잘 훈련된 운동선수에서 증가된 Scd1의 발현은 인슐린 민감성을 높이고 베타 산화 및 지질생성을 증가시킨다고 하였다. 그러나 마우스를 이용한 실험에서는 Scd1을 knock out시킨 마우스에서 인슐린 신호전달이 증가된 것이 관찰되었고 글루코스 이동 관련 GLUT4의 양이 증가된 것이 관찰되었다. 이는 본 연구에서 Scd3가 운동을 통해 발현이 줄었고 해당과정관련 유전자의 발현이 증가된 것과 맥락을 같이한다. 반면, Scd3은 마우스의 피부에서 주로 발현되고[51], 전체적인 Scd1의 억제와 간에서의 Scd3의 과발현을 유도한 마우스의 경우 정상 마우스와 비교할 때 몸무게가 증가하지 않았고 생식선과 피하의 지방세포의 무게 또한 감소된 연구를 볼 때[52] Scd3의 근육에서의 기능은 상대적으로 잘 알려지지 않았고 일회성의 운동에 의해 감소된 것을 바탕으로 근육에서의 물질대사와 관련한 scd3의 역할을 더 연구할 필요성이 있다고 사료된다.
본 연구에서 사용된 마우스의 비복근은 발바닥 굴근의 역할을 하고 지근과 속근 형태의 근육이 모두 존재하여[53] 본 microarray 분석에 적합하였다. 그러나 트레드밀 운동 시에 가자미근도 주동근으로서 작동하므로 가자미근을 이용한 추가적인 microarray 분석이 필요할 것이다. 또한 microarray 분석 기법을 사용한 연구에서는 그 목적에 따라 다양한 샘플의 수를 설정할 수 있는데 선행연구[54-56]에서는 3마리 또는 4마리로 그 수를 설정하였다. 본 연구에서는 유전형이 동일한 마우스를 대조군 3마리, 운동군 4마리를 이용하여 분석을 진행하였고 또한 Fig. 1과 같은 계층화 분석도에서 그룹 내 샘플 간 유사성이 관찰되었다. 끝으로 통계적 기준에 따라 서로 다른 기준의 차별발현 유전자들이 도출될 수 있으므로 통계적 설정 또한 고려해야 할 사항이다.
결 론
본 연구의 결과를 통해 도출된 차별발현 유전자 중 Nr4a3, Nr4a2, Btg2, Otud1, Sik1, Thbs1, Irs2가 일회성 운동 자극 후 상향조절된 상위 10개의 유전자에 속하였으며 Ube2l6, Scd3의 경우는 가장 높은 정도로 하향 조절된 유전자에 포함되었다. 또한 HK1, HK2, Adh1의 경우는 물질 대사와 관련하여 통계적으로 유의하게 관찰되었다. 따라서 일회성 운동 자극 후 가장 높게 상향 및 하향 조절된 이 유전자들을 에너지 대사 관련 생리에 역할을 할 수 있는 유전자들로 제안한다. 추후, 1시간의 간헐적인 운동의 반복실행에 의한 이들 유전자들의 변화가 대사 증후군과 관련한 신호전달이나 물질들에 영향을 끼쳐 최종적인 에너지 대사 프로파일의 개선에 도움이 될 수 있는지는 검증이 필요하다. 특히 가장 높은 수준으로 변한 Nr4a3와 NR4a2의 경우 아직 운동과 관련하여 그 영향과 기전에 관한 연구가 미비하므로 대사 적응과 관련한 운동자극 중개물질로서 지질 및 단백질 탄수화물 등의 다양한 기질 대사와의 관계성이 연구되어야 한다고 사료된다.