서 론
텔로미어는 진핵세포 염색체의 끝에 헥사뉴클레오티드 시퀀스(hexanucleotide sequences)로 구성되어 있는 특화된 DNA 캡(specialized DNA caps)으로, 재결합(recombination), 융합(fusion), 감손(degradation)으로부터 염색체 말단의 안정성을 유지하고 유전정보를 보존하며, 세포의 복제능력을 조절한다[1]. 이러한 텔로미어의 유전과 세포의 안정성(integrity) 유지는 단순한 DNA 서열(5’-(TTAGGG) n-3’)의 비코딩 직렬 반복영역과 선형 염색체의 끝을 덮은 쉘트린 단백질(shelterin protein), 그리고 텔로미어에 새로운 DNA를 추가하는 세포 역전사 효소(cellular reverse transciptase)인 텔로머라아제에 의해 이루어진다[2]. 짧거나 기능이상의 텔로미어는 종종 DNA 이중가닥 파괴(DNA double strand breaks)로 인식되어 세포주기 정지를 유발하고 세포 노화나 세포사멸을 초래하게 되는데[3,4], 텔로머라아제는 염색체 DNA 말단에 텔로미어 반복서열을 추가하여 DNA 복제에서 발생되는 손실을 보상하고[1,5], 텔로미어 연장(elongation)과 유지에 관여하므로 건강한 세포 기능에 필수적이다.
이러한 텔로머라아제 활성도는 미토콘드리아의 생산과 기능에도 필수적이다[6]. 미토콘드리아는 산화 스트레스에 민감한 기관으로 생체 반응에 필수적인 ATP를 생성할 뿐만 아니라 지방산의 베타 산화(β-oxidation), 크렙스 회로(tricarboxylic acid: Krebs cycle) 및 요소 회로(urea cycle)와 같은 다양한 대사적 경로를 통해 산화적 인산화(oxidative phosphorylation)과정을 수행한다. 이러한 과정에서 미토콘드리아는 반응성 산소종(reactive oxygen species, ROS)을 생성하게 되는데[7,8], 미토콘드리아 내부는 복귀 시스템과 히스톤이 부족하고, 미토콘드리아 DNA(mtDNA)는 전자 수송계의 활성산소 발생지와 인접해 있기 때문에 ROS에 의한 산화적 손상이 증가하고 산화손상의 표적기관이 된다[9]. 따라서 mtDNA 손상은 세포 내 산화적 손상의 지표로 제시되었다[10]. 반면, 텔로머라아제를 구성하는 역전사 효소들은 ROS 생성을 감소시켜 mtDNA를 보호하며, 세포 사멸(apoptosis)를 억제하여 미콘드리아를 보호하는 것으로 보고되었다[11,12].
그러나, 텔로머라아제의 활성은 성장기까지는 높은 활성을 유지하지만, 20대 이후부터는 급속히 감소하면서 성장기에 비하여 활성도가 낮아지고, 40세 이후부터는 활성이 나타나지 않는 사람도 35%에 달하게 된다[13]. 따라서 나이가 들어감에 따라 백혈구 텔로미어는 느리고 점진적으로 짧아지게 되고[14], 세포의 복제기능 노화와 함께 미토콘드리아의 기능변화가 나타나게 된다. 텔로미어 길이의 감소는 개인마다 많은 차이를 나타내며, 많은 선행연구들이 장기간의 규칙적인 신체활동이 텔로미어 길이 보존에 긍정적인 영향을 주고, 높은 신체활동량을 가진 사람들의 텔로미어 길이가 동일 연령의 좌업생활자보다 더 긴 것으로 보고하였다[15-18].
그러나 중·고강도의 신체활동과 백혈구 텔로미어 길이의 관련성은 대상자들의 연령에 영향을 받는다. 청소년(14-18세)[19]과 젊은 성인집단(22-27세)[20]에서는 신체활동량과 텔로미어 길이의 관련성이 나타나지 않지만, 중년집단(55-72세)[21]과 노인집단(66-77세)[20]에서는 규칙적으로 운동을 실시한 집단이 좌업생활자에 비하여 긴 것으로 보고되었다. 이러한 차이는 젊은시기에는 텔로미어의 감손이 나타나지 않는 시기이므로 20-30대의 텔로미어 길이는 신체활동량과 관련이 없지만[21], 중년 이후의 규칙적인 유산소 운동은 미토콘드리아의 밀도를 증가시키고 항산화 작용을 활성화시켜 산화스트레스를 감소시키므로 미토콘드리아 DNA 변이를 방지하여 텔로미어 길이를 유지하는 데 도움이 되는 것으로 제시되었다[22].
그러나 중정도의 신체활동량을 가진 사람들이 더 긴 텔로미어 길이를 나타내는 것과 다르게 너무 많은 신체활동량을 가진 사람들은 더 짧은 텔로미어 길이를 나타낸다는 보고[16]도 있으며, 유산소 운동과 달리 저항운동에 따른 혈액과 골격근 DNA 텔로미어 길이는 좌업생활자와 노인을 대상으로 6개월간의 저항운동 후 텔로미어 길이와 텔로머라아제의 변화가 없었다는 연구[23]와 8년 이상의 파워리프트 경력을 가진 선수들의 텔로미어 길이가 일반인보다 긴 경향이 있었으나 스쿼트와 데드리프트 기록과 역의 상관관계를 나타내었다는 보고[24] 등 저항운동과 텔로미어 길이의 관련성은 아직 일치되지 않는 보고들이 이루어지고 있다.
따라서 이 연구는 신체활동량과 텔로미어 길이가 관련성을 보이는 40-50대 중년 남성들을 대상으로 운동유형(유산소 운동과 저항운동)과 신체활동수준에 따른 텔로미어 길이의 차이를 알아보고자 한다. 이를 위하여 근력운동을 주로 수행하는 보디빌딩 종목 또는 유산소 운동을 주로 수행하는 사이클링 종목에서 최소 5년 이상 선수생활을 하였고 현재까지 규칙적으로 주당 150분 이상의 중강도 이상의 신체 활동을 수행하는 보디빌딩 및 사이클링 집단과 동일한 연령의 대조군(주당 중강도 이상의 신체활동을 150분 미만으로 수행하는 집단)에 따른 텔로미어 길이의 차이를 분석함으로써 세포 노화 예방에 미치는 운동형태의 차이를 알아보고자 한다.
연구 방법
1. 연구 대상
이 연구는 텔로미어와 운동량에 따른 관련성을 분석하기 위하여 40-50대(44.7±4.1세, 40-56세)의 남성 109명을 대상으로 실시하였다. 집단의 구분은 최소 5년 이상 보디빌딩 또는 사이클링 선수생활을 하였고 각 운동형태에서 주당 150분 이상, 중강도 이상의 신체활동을 하는 집단 중 유산소 운동집단(사이클 선수, n =37), 저항운동집단(보디빌더 선수, n =36)과 동일한 연령대의 대조군(n =36)으로 주당 중강도 이상의 신체활동을 150분 미만으로 수행하는 통제집단(CON)으로 분류하였다. 대상자는 체육협회의 웹 사이트 광고, 생활스포츠 동호회 및 SNS 홍보를 통해 모집하였다. 모든 대상자들은 측정변인에 영향을 줄 수 있는 대사장애, 항염증제나 스테로이드 또는 최근 3개월 이내에 외과적 수술 경험이 없는 자를 대상으로 하였다. 이러한 기준에 따라 참여자를 선별하기 위하여 설문지는 운동선수 경력과 운동 유형, 신체활동강도, 빈도 및 지속시간, 이전의 질병과 수술 경력, 흡연, 알코올 섭취 등 생활습관 및 병력과 관련된 질문지로 작성되었다. 이 연구는 I대학교 기관윤리위원회(No: 180309-1A)에 의해 승인된 절차에 따라 실시하였다.
2. 연구절차
1) 신체구성 측정
신체구성은 훈련된 검사자에 의해 측정되었다. 신장, 체중, 체지방률은 가벼운 옷차림을 한 상태에서 신발을 벗고 직립한 자세로 체성분분석기(Inbody 4.5, Biospace, Korea)를 사용하여 생체전기저항법(Bioelectrical impedance analysis)으로 측정하였다. 또한, 신장과 체중을 이용하여 체질량지수(BMI)는 체중(kg)/신장(m2)의 계산법에 따라 계산하였다. 허리둘레(waist circumference, WC)는 평평한 바닥에 서있는 상태에서 줄자를 이용하여 측정하였으며, 호기한 후 늑골과 장골능 사이의 가장 가는 부위를 측정하였다.
2) 신체활동 측정
이 연구에서 신체활동량을 추정하기 위해 국제합의기구에서 개발된 신체활동설문지(International Physical Activity Questionnaire, IPAQ)를 사용하였다. IPAQ는 여가시간, 실내활동, 야외활동, 일 관련, 교통 관련 활동 등을 모두 포함하는 포괄적인 범위에서 행해지는 모든 신체활동을 측정하며, 지난 7일 동안 시행한 10분 이상의 고강도( > 6 METs), 중강도(3-6 METs) 활동 및 걷기(< 3 METs) 시간을 평균 시간과 빈도를 작성하도록 되어 있다[25]. 이 실험지는 2000년에 12개국에서 신뢰도와 타당도 조사가 이루어졌으며[26], 장문형, 단문형 자기기입식 설문지와 전화를 통한 설문 방식으로 나누어진다[27]. 이 연구에서는 자기기입식 설문방식을 이용하여 신체활동량을 측정하였다.
3) 미토콘드리아 DNA 복제수와 DNA 산화 스트레스 측정
채혈은 최소 10시간의 공복상태로 하여 오점 8-10시 사이에 실시하였다. 연구대상자에게 채혈 전 24시간 이내에 고강도 운동을 하지 않도록 주의를 주었으며, 채혈 10분 전에 실험실에 도착하여 충분한 안정을 취한 후 전완주정맥에서 10 mL의 혈액을 채취하였다.
혈액에서 미토콘드리아의 DNA를 추출하기 위해 QIAamp DNA Mini Kit (The QIAamp DNA Mini Kit, QIAGEN, Chatsworth, California, USA)을 이용하였고, 각 시료의 농도는 GeneQuant 분광광도계(GENEAUANT 1,300 spectrophotometer, Biochrom, Ltd., Cambridge, USA)를 이용하여 측정하였다. 미토콘드리아 DNA (mtDNA) 농도는 실시간 중합효소 연쇄반응(a real-time polymerase chain reaction, Applied Biosystems, Foster, California, USA)을 이용하여 측정하였다. 핵 유전체 당 상대적 mtDNA 복제 수(copy number)는 69 bp fragment (nucleotides 14918-14986)와 77 bp fragment (β-globin) of mtDNA의 상대 복제 수로 계산하였다. 프라이머(primers)와 6-카르복시플루오레세인(6-carboxyfluorescein, FAM)이 표지된 Taqman 6-카르복시테드라메틸-로다민(6-carboxytetramethyl-rhodamine, TAMRA) 탐침자(probes)로 설계된 프로그램을 이용하여 2번 반복측정하였다. 중합효소 연쇄 반응을 위해 1X Taqman Universal PCR Master Mix (4304437; Applied Biosystems), 1 μM 정방향(forward)과 역방향(reverse) 프라이머 및 0.25 μM (FAM) labeled Taqman/TAMRA 탐침자를 DNA 20 ng과 혼합하였다. 중합효소연쇄반응을 위해 50°C에서 2분간 uracil-DNA-glycosylase, 95°C에서 60초간 변성(denaturation)과 효소활성화를 위해 10분을 35회 주기로 설정하였다. 95°C에서 60초 동안 변성, 60°C에서 40초 동안 primer 가열냉각, 그리고 72°C에서 30초 동안 연장 단계로 프로그램화하였다. 증폭주기의 마지막 단계는 72°C에서 7분으로 설정했다. 형광 스펙트럼(fluorescence spectra)을 통해 각 PCR 사이클의 신장단계 동안 기록된 임계주기수(the threshold cycle number, Ct)를 기록하였고 다음의 공식을 사용하여 계산하였다(Rc =2ΔCtΔCt = Ct β-globin-Ct mtDNA).
8-OHdG는 DNA 산화 지표로 분석하였다. 8-OHdG는 면역분석(Immmunoassay) EIA kit (DNA/RNA oxidative damage ELISA kit, Cayman Chemical, Michigan, USA)을 이용하여 anti-mouse IgG-coated plate와 8-OH-dG-enzyme conjugate로 구성된 추적체(tracer)를 사용하여 제한된 양의 8-OHdG의 분석을 위해 8-hydroxy-2-deoxy guanosine과 8-OH-dG acetycholinesterase (AChE) conjugate을 이용하여 분석하였다. 분석방법은 매뉴얼에 따라 수행하였으며, 실험방법은 다음과 같다. 8-OHdG 복합체(complex) 항체를 벽(well)에 고정시켜 둔 goat polyclonal anti-mouse IgG에 결합시키고 결합되지 않는 반응물을 제거하기 위해 플레이트(plate)를 씻은 후 Ellman's Reagent (AChE)를 벽에 첨가하였다. 효소 반응에 의한 생성물은 표준비색 UV 분광 광도분석(a standard colorimetric UV spectrophotometric assay, ThermoFisher Scientific, Fredrick, MD, USA)을 이용하여 412 nm에서 관찰하였으며, 8-OHdG 농도는 8-OHdG 표준곡선(standard curve)을 이용하여 결정하였다.
4) 텔로머라아제 활성도 측정
텔로머라제 효소 활성도는 telomere repeat amplification protocol의 kit (TRAPeze Telomerase detection kit, Chemicon, NY, USA)을 이용하여 실험절차에 따라 분석하였다. 분석절차는 kit의 매뉴얼에 따라 분석하였으며, 텔로머라아제의 활성도는 표준비색 UV 분광 광도분석(ThermoFisher Scientific, Fredrick, MD, USA)을 이용하여 620 nm에서 관찰된 값으로 분석하였다.
5) 텔로미어 길이 분석
텔로미어 길이를 측정하기 위하여 Wizard Genomic DNA Purification Kit (Promega, Medison, USA) 프로토콜을 사용하여 말초 혈액으로부터 DNA를 추출하였다. 텔로미어 복제 수(copy number)에 대한 단일 유전자 복제 수의 상대적 비는 Cawthon et al. [28]에 의해 기술된 정량적 RCR (quantitative PCR) 측정방법을 이용하여 Real-Time PCR System (Applied Biosystems, Foster, California, USA)을 이용하여 분석하였고, 상대 텔로미어(T)/단일유전자(S)를 측정하기 위해 표준 곡선을 이용하여 텔로미어 길이를 제시하였다. 텔로미어 길이 분석방법은 다음과 같다. 텔로미어 primer(5→3')는 tel 1, GGTTTTTGAGGGTGAGGGTGAGGGTGAGGGTGAGGGT; tel 2, TCCCGACTATCCCTATCC CTATCCCTATCCCTATCCCTA를 사용하였다. 단일 유전자 primer는 36b4u, CAG-CAAGT GGGAAGGTGTAATCC; 36B4d. CCCATTCTATCATCAACGGG TACAA를 사용하였다. 중합 효소 연쇄반응을 위한 텔로미어 primer는 tel 1, 270 nM; tel 2, 900 nM를 혼합하였고, 단일 유전자 primer로 사용된 36B4 primer 농도는 36B4u, 300 nM; 36B4d, 500 nM를 혼합하였다. 중합 효소 연쇄 반응을 수행하기 위해 94°C에서 3분 동안 변성을 시작했고, 다음과 같은 35회 주기로 이루어졌다: 94°C에서 30초 동안 변성, 56°C에서 25초 동안 primer 가열냉각, 그리고 72°C에서 30초 동안 연장 단계로 프로그램화 하였다. 증폭주기의 마지막 단계는 72°C에서 7분으로 설정했다.
3. 자료처리방법
본 연구의 자료 분석을 위해 SPSS WIN 23.0 프로그램을 사용하였으며, 측정요인들에 대한 평균과 표준편차를 산출하였다. 집단 간 신체 구성, 신체활동량, mtDNA 복제 수, DNA 산화 스트레스, 텔로머라아제 활성도와 텔로미어 길이는 일원변량분석(One-way ANOVA)을 이용하여 분석하였다. 신체활동량, mtDNA 복제 수, DNA 산화 스트레스, 텔로머라아제 활성도와 텔로미어 길이의 관련성은 피어슨 상관계 수(Pearson correlation)를 이용하여 분석하였다. 사후검증은 Scheffe를 이용하였으며, 모두 유의수준 α=.05에서 검증하였다.
연구 결과
1. 운동유형에 따른 신체구성과 신체활동량
나이(p< .001), 체중(p< .001), 체질량지수(p< .01), 근육량(p< .001), 체지방률(p< .001), 고·중강도 신체활동량(p< .01)이 집단 간 유의한 차이가 나타났다(Table 1). 사후검증 결과 나이는 BB 집단이 CC와 CON 집단에 비하여 유의하게 낮은 것으로 나타났다(p< .001). 체중(p< .001)과 체질량지수(p< .01)는 BB 집단이 CC 집단보다 유의하게 높았으며(p< .001), 근육량은 BB 집단이 CC와 CON 집단에 비하여 유의하게 높은 것으로 나타났다(p < .001). 고·중강도 신체활동량은 BB와 CC집단이 CON 집단에 비하여 유의하게 높은 것으로 나타났으며(p< .05), BB와 CC 집단의 비교에서도 BB 집단이 유의하게 높았다(p< .05).
2. 운동형태에 따른 mtDNA 복제 수, DNA 산화 스트레스, 텔로머라아제와 텔로미어 길이
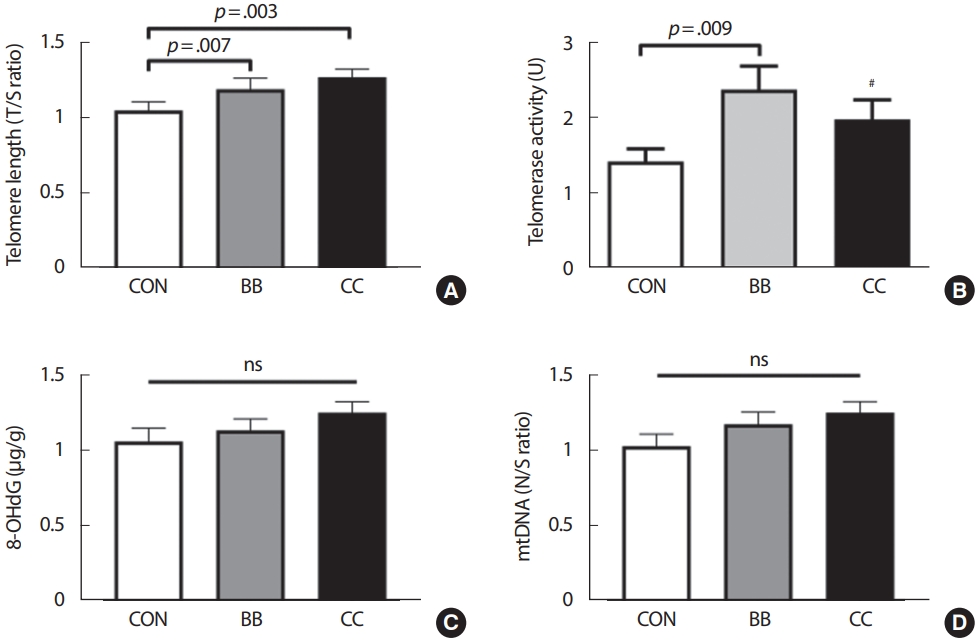
텔로머라아제(p< .05)와 텔로미어 길이(p< .01)는 운동 형태 또는 종목에 따른 유의한 차이를 나타내었다(Fig. 1). 사후검증 결과 텔로머라아제는 BB 집단이 CON 집단보다 유의하게 높은 활성도를 나타냈으나(p < .05) CC집단의 경우는 높은 경향만 나타났다(p =.115). 이에 CON 집단과 CC 집단 간의 실제적 유의성(practical significance)을 확인하기 위하여 Cohen’s d 효과 크기를 산출한 결과 0.42891라는 의미 있는 중간 정도의 유의성을 확인할 수 있었다. 텔로미어 길이는 BB(p< .05)와 CC (p< .01)집단이 CON 집단보다 유의하게 긴 텔로미어 길이를 나타냈다. 한편 8-OHdG와 mtDNA는 집단 간 유의한 차이가 나타나지 않았다.
3. 신체활동량, 신체조성 및 텔로미어 관련 변인들 간의 상관성
고·중강도 신체활동량은 나이(r= -.386, p< .001), 체지방률(r= -.418, p< .001)과 유의한 음의 관련성을 나타냈으며, 근육량(r=.303, p< .01)과 유의한 양의 관련성을 나타냈다. 고·중강도 신체활동량은 텔로머라아제 활성도(r=.266, p< .01), mtDNA 복제 수(r=.195, p< .05) 및 텔로미어 길이(r=.285, p< .01)와 유의한 양의 관련성을 나타냈다. 텔로머라아제는 텔로미어 길이(r=.625, p< .001), mtDNA 복제 수(r=.519, p< .001)와 유의한 양의 관련성을 나타냈다. mtDNA 복제 수는 텔로미어 길이(r= .771, p< .001)와 유의한 양의 관련성을 나타냈다(Fig. 2).
논 의
이 연구에서 텔로미어 길이는 운동형태와 무관하게 운동량에 따른 차이를 나타냈으며, 주당 150분 이상의 고·중강도 신체활동량을 가진 BB와 CC 집단이 CON 집단에 비해 더 긴 텔로미어 길이를 가지고 있는 것으로 나타났다(p< .01). 반면에 텔로머라아제 활성도는 CON 집단에 비해 BB 집단은 유의하게 증가하였으나 CC 집단에서는 높은 경향만(p =.115)을 보였다. 하지만 실제적인 유의성을 대변하는 Cohen’s d 효과 크기(d =.42891)가 의미 있는 수준에서 나타남을 확인할 수 있었다. 운동형태에 따른 텔로미어 길이에 대한 연구들은 대부분의 연구에서 지구력 또는 유산소성 운동이 텔로미어 길이와 양의 관련성을 나타내는 것으로 보고하였으나[15,18,21], 장기간의 저항훈련에 대한 연구는 매우 부족한 실정이다. Kadi et al. [24]의 연구에 따르면 평균 7년 이상의 역도 선수 경험을 하고 8년 이상 역도 훈련에 참여한 젊은 남성 선수들이 활동적인 동 연령대 남성에 비해 통계적으로 유의하지는 않았으나 골격근의 텔로미어 길이가 긴 경향을 나타냈다고 보고하였다. 또한 6,503명의 성인을 대상으로 실시한 횡단연구에서는 중강도 이상의 유산소 신체활동과 근력운동을 병행한 경우, 신체활동량이 많을수록 백혈구 텔로미어 길이가 유의하게 긴 것으로 나타났다[29]. 반면, Werner et al. [30]의 연구에서는 각기 다른 운동 형태에의 6개월 동안의 훈련이 백혈구 텔로미어 길이 변화에 미치는 영향을 검토하였고 그 결과로 저항운동과는 달리 유산소 운동 훈련에서만 유의하게 긴 것으로 나타났다. 이러한 연구 결과는 본 연구결과와는 상반되는 것이다.
일반적으로 중강도 이상의 유산소 신체활동은 텔로머라아제의 활성을 증가시키고 이러한 활성증가는 림프구(lymphocyte), 과립구(granulocytes) 및 백혈구(leucocytes)의 텔로미어 길이를 증가 또는 감손을 억제하는 것으로 보고되고 있다[31]. 하지만 텔로머라아제 활성을 증가시켜 텔로미어 길이를 증가시키거나 감손을 억제하는 잠재적인 매개체로는 IGF-1과 혈관 전단 스트레스(vascular shear stress)로 나타나는 산화질소(nitric oxide, NO)가 제안되고 있다. 본 연구에서는 Werner et al. [30]의 연구와는 달리 저항성 운동을 주로 하는 보디빌딩 집단에서 텔로머라제의 활성 증가뿐만 아니라 텔로미어 길이 역시 유의하게 긴 것으로 나타났다. 이러한 차이는 아마도 수행하는 운동강도와 운동량의 차이에서 비롯될 수 있다. Werner et al. [30]의 연구에서는 8가지 항목을 20-RM의 강도 즉, 약 50% 1-RM의 강도로 주당 3회, 1회 45분(45×3 =135분)간 실시하는 프로토콜을 6개월간 실시한 반면 본 연구에서는 최소 5년 이상, 평균 10년 정도를 보디빌더 선수로 활동한 남성들로 근비대를 위한 강도는 일반적으로 70% 이상의 고강도로 저항운동을 실시할 뿐만 아니라 주당 신체활동량이 평균 494.9±323.6 (분)으로 3배 이상 차이가 난다는 점이다. IGF-1은 훈련되지 않은 남성에 비해 장 기간의 저항훈련을 수행하는 사람들에서 안정 시 IGF-1의 농도가 높은 것으로 보고되었으며[32], 특히 훈련량(high volume, multiple set)과 운동강도가 안정 시 IGF-1 농도 증가에 중요한 요소로 작용하기에[33,34] 선행연구와 차이가 나타날 수 있다. 즉, 이 연구에서의 보디빌딩 집단(BB, 저항운동)의 텔로머라아제의 활성 증가 및 텔로미어 길이가 CON 집단에 비해 더 긴 것은 아마도 장기간의 저항운동이 일시적 또는 장기적인 IGF-1의 증가를 유발하고 이를 매개로 나타난 것으로 판단된다. 추후 연구에서는 이에 대한 확인 작업이 필요할 것으로 사료된다.
이 연구에서 운동종목으로 분류하여 분석한 결과, 텔로미어 길이는 운동형태와 관계없이 운동선수집단이 대조군보다 더 긴 텔로미어 길이를 가진 것으로 나타났다. 반면, 고·중강도 신체활동량은 관련성이 약하기는 하지만 텔로미어 길이(r=.285, p< .01)와 유의한 양의 관련성을 나타내었다. 이러한 결과는 운동종목이나 형태와 관련 없이 장기간의 고·중강도의 신체활동이 텔로미어 길이와 관련이 있을 것으로 추측해 볼 수 있다.
이 연구에서 유산소성 운동집단보다 저항운동 집단에서 CON 집단보다 더 높은 텔로머라아제 활성을 나타낸 결과는 흥미롭다. 저항운동과는 달리 지구성 운동은 혈관 전단 스트레스를 증가시킴으로써 그 결과물로 산화질소를 방출한다[35]. 특히, 내피의 산화질소 합성과 텔로머라아제 활성도는 운동에 의한 혈관 보호 신호경로(signaling pathway)와 서로 연관되며[36], 혈관 내피세포에서의 NO 합성을 통하여 혈관 노화를 방지하는 것은 텔로머라아제 활성 및 텔로미어 길이 증가와 관련된 것으로 보고되고 있다[36]. 따라서 저항성 운동은 IGF-1의 매개를 통해 그리고 유산소 운동은 혈관 내피 세포의 NO의 합성 증가를 통해 텔로머라아제의 활성을 증가시켜 텔로미어의 길이 감손을 억제할 수 있을 것으로 판단된다.
그러나 많은 양의 고강도 저항운동을 실시하는 선수들은 운동 후 근통과 근경직 등과 같은 fatigue athlete myopathic syndrome (FAMS)을 나타내며, 이러한 증상을 가진 선수들의 외측광근 텔로미어 길이가 일반인에 비하여 짧은 것으로 보고되었다[37]. 또한 신체활동량과 텔로미어 길이와의 U 곡선(U curve)를 나타내는 것으로 보고되는[17] 등 지나친 신체활동량이 텔로미어 길이 감손에 부정적인 영향을 준다는 연구들도 보고되었다. 그 이외에도 일회성 고강도와 고운동량의 신체활동은 일시적으로 텔로미어 DNA 부위에 산화적 스트레스를 주어 손상을 줄 수 있다는 연구들이 보고되고 있다[38]. 이러한 선행연구들의 보고는 이 연구와는 상반된 결과라 판단된다. 특히 이 연구에 참여한 BB와 CC 집단의 대상자들은 평균 10년가량의 오랜 선수 경력을 지닌 중년 남성들로 고강도와 많은 신체활동량의 훈련 경력을 지닌 대상자들로 구성되었기 때문이다. 이러한 선행연구와의 차이는 다양한 원인(집단의 동질성, 샘플 사이즈, 연령대, 성별 등)으로 설명될 수 있지만 아마도 첫째, 일시적 반응과 장기간의 적응 차이이다. Borghini et al. [38]의 연구에서와 같이 17명의 40대 지구성 남성 선수를 대상으로 혈중 텔로미어 길이를 동 연령대의 좌업자와 비교했을 때 유의하게 길었음을 보고한 반면 이들을 대상으로 울트라 마라톤을 수행한 후 측정한 결과에서는 유의하게 짧아졌음을 보고한 바 있다. 즉, 장기간의 운동을 통한 텔로머라아제 활성의 상향 조절은 산화적 스트레스에 대한 항산화 방어[39], DNA 복구 효소의 증가[40], ROS의 생성 감소[41]를 통해 이루어진다. 이 연구에서 DNA 손상지표로 측정된 8-OHdG 수준과 mtDNA 복제 수의 집단 간 차이가 없었던 결과를 고려해보면, 운동형태와 관계없이 장기간의 고·중강도 신체활동량이 고강도 운동으로 유발될 수 있는 산화스트레스에 대한 방어시스템을 강화하거나 적응력을 증가시켜 텔로머라아제 활성과 텔로미어 길이에 영향을 주었을 가능성이 있다. 둘째, 어떤 부위(근육 vs. 혈중)의 텔로미어 길이를 관찰하느냐의 차이로 기인할 수 있다. 즉, 혈중 마커는 전신을 반영할 수 있으나 특정 부위의 근육에서의 텔로미어 길이 분석은 국소적인 경향이 있으며 특히 어떤 자극 조건이냐에 따라 결과가 달라질 수 있다. Kadi et al. [24]의 연구에서는 8년 정도의 역도선수 경력을 지닌 남성 선수(28.5±6.6세)와 동연령대의 활동적인 남성의 외측광근 텔로미어 길이를 비교한 결과, 역도 선수들의 텔로미어 길이가 긴 경향(p=.07)을 보였으나 스쿼트와 데드리프트 기록과는 역상관을 지닌다는 결과를 보고한 바 있다.
셋쩨, 장기간의 규칙적인 운동에 따른 신체조성의 변화이다. 과도한 체지방은 전신염증, 산화스트레스 및 백혈구의 교체를 증가시켜 텔로미어 길이 감손을 촉진시키는 것으로 보고된 바 있다[42]. 그러나 이 연구에서 CC 집단은 BB 집단에 비하여 유의하게 낮은 BMI와 체지방률을 나타내었고(p< .001), BB 집단은 CC와 CON 집단에 비하여 유의하게 높은 근육량을 나타냈다(p< .001). 또한 운동량은 근육량과는 유의한 양의 관련성(p =.001)을 나타내고 체지방률과는 유의한 음의 관련성(p< .001)을 나타냈다. 따라서 장기간의 운동에 따라 적절한 수준의 체지방과 근육량을 중년에도 유지하게 하고, 이러한 신체구성이 염증과 산화스트레스를 어느 정도 억제하여 텔로미어 길이에 영향을 주었을 가능성이 있다.
넷째, 고·중강도 신체활동량이 텔로머라아제 활성도(r=.266, p< .01), mtDNA 복제 수(r=.195, p< .05) 및 텔로미어 길이(r=.285, p< .01)와 유의한 양의 관련성을 나타냈고, 이러한 결과는 관련성이 낮은 수준이지만, 중년 남성에서의 평균 10년 이상의 장기간의 고·중강도 신체활동이 텔로미어 길이에 영향을 주었을 가능성이다. Saßenroth et al [43]은 현재 비활동 1-9년 동안의 신체적 활동기간, 10-41년 동안의 신체적 활동기간 그리고 41년 이상의 신체적 활동기간으로 분류하여 텔로미어와의 관련성을 살펴본 결과, 최소 10년 이상의 규칙적인 스포츠 활동이 텔로미어 길이에 영향을 준다고 보고하였다. Ludlow et al. [17]도 짧은 기간의 운동은 텔로미어 길이에 영향을 줄만큼 충분한 자극을 주지 못하지만, 5년 이상의 규칙적인 운동이 텔로미어 길이를 보호하는 효과가 있는 것으로 보고하였다. 따라서, 장기간의 운동과 중년의 운동이 텔로미어에 영향을 주는 원인을 젊은 시절(20-30세) 동안 만의 운동으로는 예측을 하기 어렵지만, 이후의 중년과 후기의 생활양식은 일상생활의 “세포 기억(cell memory)”으로 남아 긴 텔로미어를 가진 세포의 복제를 통한 재생산 혹은 분열(mitosis)하기 전 세포의 유입과 같은 기전의 변화를 가져와 텔로미어 길이에 영향을 주었을 가능성이 있다고 제안하였다[36]. 따라서 운동종목 또는 형태에 따른 차이보다는고·중강도의 지속적인 신체활동이 텔로머라아제 활성도를 높게 유지시키고 텔로미어 길이 감손을 억제한 것으로 판단된다.
이 연구의 제한점은 세포노화 방지를 위한 적정 운동량의 기준을 제시하지 못한 점이다. 이는 신체활동량 측정이 자기-보고식 설문지를 사용하였기 때문에 현재의 건강상태와 컨디션 및 주관적인 운동자각도에 의해 운동강도가 작성되고 사회적인 기대감에 따라 운동량이 객관적인 기구를 이용하여 측정된 운동량보다 과다하게 보고된다는 것이다[44]. 이 연구에서도 운동집단의 경우 신체활동량이 건강을 위한 신체활동량 지침보다 높게 나타나 보편적으로 제시하고 적용하기에는 적절하지 않은 것으로 판단되었다. 또한, 운동에 반응하는 다양한 면역인자와 호르몬들에 대한 측정이 이루어지지 못하여 장기간의 중-고강도 운동이 텔로미어 길이 감손에 긍정적인 영향을 주는 기전에 대한 설명이 명확히 이루어지지 못하였다. 따라서 이와 관련된 후속 연구들이 이루어져야 할 것으로 제시하는 바이다.
결 론
이 연구에서 평균 10년 이상의 선수생활을 하였고 최근 3개월 이상 주당 150분 이상의 고·중강도 신체활동을 하는 BB 집단과 CC 집단 그리고 주당 150분 이하의 고·중강도 신체활동을 수행하는 CON 집단을 비교한 결과, 운동 종목이나 형태와 관련없이 고·중강도의 높은 신체활동량을 수행하는 집단이 텔로머라아제의 활성을 증가시켜 텔로미어의 감손을 예방하는 것으로 나타났다. 특히 이 연구는 중년 남성들을 대상으로 40-50대의 연령을 한정하여 실시하였기 때문에 중년 남성에서의 지속적인 고·중강도 신체활동의 노화방지에 대한 효과를 나타낸 의의가 있는 연구로 생각된다.