


INTRODUCTION
Large-volume skeletal muscle loss usually results in functional deficits, cosmetic flaws, emotional pain, and a permanent disability [1]. Volumetric muscle loss (VML) occurs due to motor vehicle accidents and surgical ablation, including tumor excision and diabetic foot debridement in civilians and gunshots or explosive-related injuries in military personnel. Severe full-thickness muscle loss creates a large gap between the end of the transected muscle stumps that hampers the natural regenerative process and consequently triggers fibrosis [2], which obstructs myofiber ingrowth and blood vessel infiltration because of the existing connective tissue structure. As a VML injury results in muscle loss and damages to nerves, blood vessels, and the extracellular matrix (ECM), the completion of functional and morphological repair remains a big challenge. Therefore, the treatment of VML injuries should simultaneously target enhanced myogenesis and inhibition of fibrosis.
Appropriate scaffolding provides a space for the ingrowth of new myofibers which can inosculate with the remaining myofibers. Biological scaffold transplantation using myogenic cells is a research hotspot in regenerative medicine. Surgical free-muscle transplantation is a standard treatment for VML and often causes not only donor-site morbidity and functional deficits, but also is unsuitable for areas with large-volume muscle tissue removal [3,4]. Thus, the transplantation of synthetic scaffolds constitutes an attractive approach because of the high reproducibility and availability despite adverse effects, including excessive inflammatory response and failure to incorporate into host tissues [5].
Alternatively, acellular ECM can facilitate tissue repair in tissues, such as cardiac muscles, skeletal muscles, and the abdominal wall [6,7]. Acellular ECM preparation involves several processes, such as tissue harvesting, decellularization, and sterilization. Nonetheless, the residual small amount of DNA in acellular ECM confers a risk of immunological rejection of xenogenic scaffolds. However, ECM component proteins are seemingly highly conserved across species [8]. In a clinical study, the transplantation of acellular porcine ECM to patients with skeletal muscle injury of the quadriceps resulted in the formation of new muscle tissues after 36 weeks post-transplantation and even evoked remarkable contractile properties [9].
Compared with no treatment, there is emerging evidence that diverse treatments/remedies for VML can ameliorate muscle recovery and function. However, the favorable impact of the therapies on muscle regeneration from VML is uncertain. Thus, novel therapeutic treatment paradigms must be developed to provide clinically appreciable changes in muscle functional capacity. This brief review summarizes the clinical outcome of acellular biomaterials or biological ECM in the absence or presence of satellites cells, progenitor cells, and growth factors that enhance functional capability after VML damage.
METHODS
Using online databases such as Pubmed, Web of Science, Google Scholar, and Wiley Online Library, previous studies on the VML and potential therapeutic interventions were collected and analyzed for this review study. The web databases were searched using terms including “ satellite cells”, “ skeletal muscle regeneration”, “ VML injury”, “ ECM scaffold”, “ resistance training”, “ angiogenesis”, and “ innervation” for data collection.
RESULTS
1. ECM as a biological scaffold
The ECM is a three-dimensional structure that consists of glycoproteins, heparin sulfate proteoglycans, glycosaminoglycans, and type IV collagen, all of which play a crucial role in skeletal muscles function and structural support [10]. The ECM communicates with cells through integrin subunits to regulate the activation, proliferation, and differentiation of progenitor cells [11]. The activity of myogenic cells depends heavily on ECM components, especially primary ECM components such as collagen, laminin, and fibronectin, have been investigated extensively. Agarose hydrogel scaffolds coupled with laminin promotes neurite extension from the dorsal root ganglia [12]. Bioengineered hydrogel, when used with collagen and fibrin, increase functional properties and induce cellular hypertrophy [13]. In contrast, the inhibition of collagen synthesis hampers myoblast differentiation [14]. Myogenic cells that are cultured on dishes coated with ECM component proteins promote myoblast proliferation, and differentiation, especially with regard to satellite cells (SC) that adhere to laminin and increase motility [15–17]. Duchenne’ s muscular dystrophy (DMD), which is characterized by an inability to produce dystrophin that makes the sarcolemma brittle, illustrates the importance of ECM proteins in muscle structure and function. Patients with DMD experience progressive muscle degeneration that culminates in premature death.
ECM elasticity is one of the factors that determine stem cell lineage specification. Mesenchymal stem cells (MSCs) cultured on collagen-coated plates with different elasticity, to mimic the stiffness of the original tissues, differentiate into neural tissues on a soft plate, into myogenic tissues on a medium-stiff plate, and into bones on a rigid plate [18]. MSCs cultured on a stiff surface express smooth muscle cell markers, whereas MSCs cultured on a soft surface express chondrogenic markers. Moreover, stem cell proliferation depends on tissue stiffness. The ratio of MSC proliferation in response to transforming growth factor-β (TGF-β) depends on the matrix stiffness [19].
Furthermore, the ECM serves as a reserve of growth factors and binds to and releases growth factors [20], including fibroblast growth factor (FGF), TGF-β, hepatocyte growth factor (HGF), and VEGF. During the repair phase of muscle regeneration, chemokines that are released from ECM degradation attract progenitor cells and stem cells [21,22]. After injures, macrophages and SCs release matrix metalloproteinases (MMPs) that promote the secretion of growth factors which bind to the ECM. The ECM-secreted growth factors enhance progenitor cell migration and proliferation [20,23,24]. Cultured SCs under high TGF-β1 signaling inhibit the MyoD and myogenin expression resulting in limited SC proliferation and differentiation [25]. Single myofiber studies show that HGF and FGF-2 affect SC proliferation [26].
Acellular ECM has significant benefits as a scaffold because antigens, which can trigger an immune response, are removed, and tissue remodeling macrophages are promoted by acellular ECM transplantation [27]. Another benefit of acellular ECM is biocompatibility and biodegradability, which induces host cells to self-produce matrix [28]. Furthermore, the 3D structure of ECM retains the original structure of myofibers, vessels, and nerves, which facilitates myotube alignment and incorporates nerves and blood vessels into the ECM [28]. Acellular ECM scaffolds seeded with progenitor cells have been extensively researched in VML injuries [28,29]. Blood vessels and myofibers have been displayed in an implanted homologous muscle acellular matrix [30]. The implantation of acellular ECM along with myoblasts can repair abdominal wall defects [31], whereas only homologous acellular ECM implantation resulted in a large volume of fibrous scar tissues [32]. Previous studies have shown the ability of the homologous acellular ECM to support muscle and blood vessel regeneration [33] where implanting an acellular ECM seeded with MSCs improves functional recovery as well [29].
2. Satellite cells as sources of muscle regeneration
SCs, which are adult stem cells, are primary resources for skeletal muscle regeneration. SCs that are located between the basal lamina and sarcolemma are mitotically quiescent. A hallmark of SC is the self-renewing capacity that has been demonstrated in SCs transplantation studies. Transplantation of SCs into SC-depleted MDX mice with a mutation in the mouse dystrophin gene induced myofiber formation, migration to the host SC niche, and transplanted SCs were are incorporation with host fibers [34]. The number of SCs in a single fiber is associated with age and fiber types, and SCs are unevenly distributed along with fiber. 30-35% of nuclei in a postnatal mouse express SC markers while an adult mouse consists only 1-4% of SC [35]. Soleus, a slow type of muscle, has two- to three-fold higher number of SCs than extensor digitorum longus, a fast type of muscle [36,37]. The population of SC is heterogenous in mammalians such as mice, rats, and humans [38–40]. A high density of SC has been observed adjacent to capillaries [41] and close to the motor neuron junction [42], which implies that they contribute to SC character. SCs are characterized by distinct markers in each sequential step. Quiescent SCs are characterized by Pax7, paired box7, which are crucial in SC development and lineage determination. Pax7- null mice appear 50% decrease in body weight at 7 days of age, and even pax7-null mice die within 2 weeks after birth [43,44].
Growth factors play a major role in SC proliferation and differentiation. Insulin-like growth factor-1 (IGF-1) is a well-known growth factor for skeletal muscle regeneration and can change the expression of myogenic regulatory factors and upregulate SC proliferation [45,46]. HGF leads SC migration to the site of damage [47] and SC proliferation by binding to c-met [48]. SCs exposed to muscle injury sites start proliferating by multiple cellular and molecular responses governing the SC environments during muscle regeneration processes. Activated SCs, often referred to as myoblasts, are characterized by the upregulation of Myf5 and myoblast determination protein 1 (MyoD) [49–51]. MyoD upregulation relates to SC proliferation, and SC differentiation depends on MyoD downregulation. MyoD-null mice have shown a limited regenerative capacity characterized by abnormal myoblasts morphology [52] and decreased the number of regenerating myofibers [53]. When SCs enter the differentiation phase, both myogenin and Myf 6 are highly expressed [54,55]. Myogenin is essential for the fusion of myoblasts, either among themselves or with existing myofibers. These small myofibers become mature fibers as they grow in size and express contractile proteins, such as actin and myosin. In contrast, myogenin-null mice have only a few myofibers at birth, and they die prenatally [56–58].
The probability of SC transplantation for muscle regenerative medicine has been extensively studied. Donor cells-derived myofibers are investigated by donor gene expression in host tissues [59,60]. Donor SCs not only fuse with themselves or host myofibers, but also exist as SCs within regenerating myofibers through self-renewal [34,61].
3. Myofibers as a source of satellite cell
SCs are the primary sources for skeletal muscle regeneration. However, enzymatically dissociated SC transplantation has the challenge of resolving skeletal muscle wasting disorders [62,63]. Enzymatically dissociated and transplanted SCs induce incomplete muscle regeneration due to limited migration of SC, donor–host histocompatibility, and early death of donor SCs. Individual, freshly isolated single myofiber transplantation is an alternative strategy to deliver SCs to damaged muscles and maintains SC stemness because SCs are influenced by the original environment, thereby increasing the survival ratio of donor SCs. Individual myofiber contains 7-22 SCs in a mouse [44,51], and SCs migrate from the fiber to the damaged site. Single myofiber containing 7 SCs generated more than 100 new fibers containing 25,000-30,000 myonuclei [59]. A single donor myofiber promotes muscle hypertrophy and an increase in cross sectional area (CSA) [64]. Single myofiber transplantation results in hypertrophy with high force production, and donor fiber-derived SCs in young age remain in a host muscle permanently, which prevents age-related mass reduction [65]. In a previous study in our labo-ratory, transplanting 50 myofibers into preembedded ECM increased the number of regenerating fibers and decreased decellularized areas in a VML rat.
4. Resistance training and muscle regeneration
Resistance training (RT) influences functional properties of skeletal muscles by modifying fiber structure, mass, metabolism and promoting the release of growth factors. The intensity and mode of exercise induce specific adaptation of skeletal muscles. RT potentially increases muscle mass and contractile properties. After 6 weeks of heavy RT, muscle strength increases by 15% [66]. RT enhances the muscle protein synthesis ratio in humans by 50% at 4 hours after heavy resistance training [67]. RT contributes to myofiber hypertrophy and skeletal muscle regenerative capacity [68,69]. SC increases by 17% after extreme 16-week RT [70]. Powerlifters with a higher SC volume respond to injuries acutely [71].
RT is one of the best treatments for VML injury because VML results in severe functional deficits. 10% of the mass of the muscle loss results in 30% of initial peak isometric force reduction [72]. When 20% of the mass of the tibialis anterior (TA) muscle is excised, the TA exhibits 29% and 31% of a functional deficit in 2 and 4 months post-injury [73]. Therefore, VML patients require special treatment to enhance functional recovery. Low-intensity exercise is beneficial to repair the cardiac tissues [74], and high-intensity exercise attracts circulating mesenchymal stem cells into the damaged tissues in myocardial ischemia patients [75]. Both resistance and endurance training trigger angiogenesis and vascularization [76,77]. It is particularly important to have vascularization when repairing or regenerating tissues to attract circulating and residence cells. Blood vessels provide nutrients and oxygen to support muscle reconstruction [78]. Collectively, as aforementioned, therapeutic effects of donor SC and resistance exercise on skeletal muscle regeneration from VML are depicted in Fig. 1.
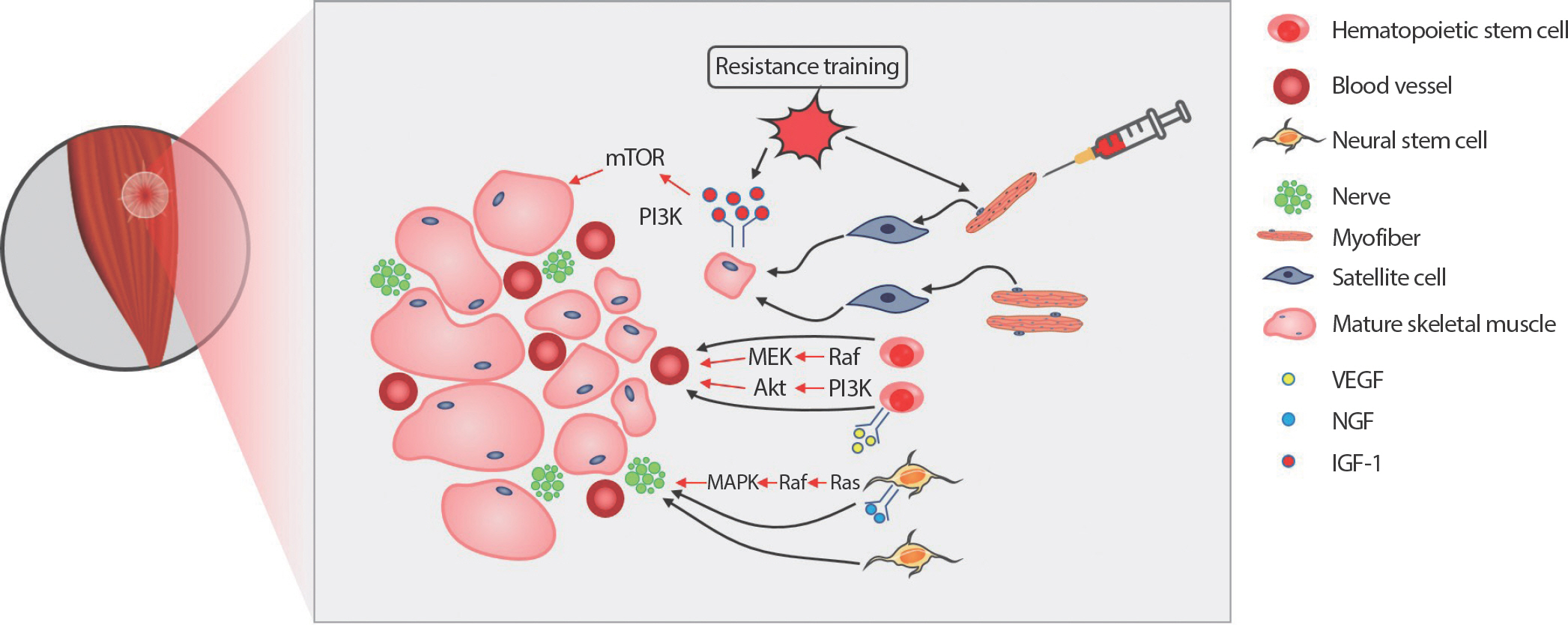
Fig. 1.
Fig. 1.Effect of resistance exercise and donor satellite cell (SC) on skeletal muscle regeneration from volumetric muscle loss (VML). Transplanted extracellular matrix (ECM) fills the gap and roles as a scaffold to infiltrate circulating cells for skeletal muscle regeneration. Resistance exercise activates SC from the single donor myofibers. Resistance exercise also stimulates insulin-like growth factor-1 (IGF-1) downstream signaling pathways, accelerating protein synthesis, and preventing protein degradation. The upregulation of the IGF-1 downstream pathways contributes to the maturation of recently regenerated small muscle fibers. Vascular endothelial growth factor (VEGF) signaling pathway initiates hematopoietic stem cells proliferation and differentiation process into angiogenic cells, which is critical for vasculogenesis and angiogenesis. Nerve growth factor (NGF) binds to tropomyosin receptor kinase A (TrkA) and actives Ras-mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K)-Akt signaling pathways to promote the survival and differentiation of neural stem cells. Successful innervation and vascularization are crucial steps for functional and structural completion of skeletal muscle regeneration in VML.
5. Innervation and vascularization of the ECM
New myofiber formation should synchronize with vascularization and innervation together because of the role of blood vessels and nerves in skeletal muscles. Transmission of nerve impulses causes skeletal muscle contraction. A nerve fiber consists of axons, and myelin sheath develops central and peripheral nerves. Motor and sensory nerve fibers involving peripheral nerves directly regulate muscle contraction. Innervation plays a vital role in functional morphological skeletal muscle maintenance [79]. Denervated skeletal muscles continue the path of degeneration due to the failure of neuromuscular junctions, resulting in atrophy and tissue necrosis [80]. Skeletal muscle regeneration occurs following denervation, but this process cannot continue when SCs are depleted [79]. As denervation results in adverse effects on the muscle, regenerating myofibers should accompany innervation in a VML injury. Schwann cells, the primary glia of the peripheral nerve fiber, induce nerve regeneration and muscle innervation.
Activated Schwann cells secrete cytokines and neurotrophic factors that guide axon growth [81]. Innervation by axons depends on the remaining Schwann cell tubes and Schwann cell extension processes leading to end plates [82]. The presence of motor neurons affects Schwann cell viability. Motor neurons secret neuregulins, which are glial growth factors that promote survival and proliferation of Schwann cells [83,84]. The effects of neuregulin on muscle through binding tyrosine kinase receptors have been observed. Human myoblasts cultured with neuregulin express an increase in acetylcholine receptors and myosin heavy chains [85]. Other studies have researched the role of neurotrophic factors such as nerve growth factor (NGF), neurotrophin-3 (NT-3), and brain-derived neurotrophic factor (BDNF) on skeletal muscle regeneration. NGF is necessary for the survival and growth of neurons. Phenotypic characterization of NGF knockout mice increases in cell death and muscle dystrophy [86]. NGF treated with laminin and muscle-derived stem cells enhance neurite extension and engraftment efficiency, respectively [12,87]. NT-3, which is a neurotrophic factor that encourages nerve growth, modulates neurogenesis. The NT-3-overexpressing transgenic mice increase sensory and motor neurons, and NT-3 delivery enhances axonal regeneration in denervated muscle [88]. BDNF supports the survival of existing neurons and promotes the growth and differentiation of new neurons [89,90]. BDNF also influences skeletal muscle regeneration. BDNF depleted mice display a decrease in Pax7 positive cells, as well as impair skeletal muscle regeneration. These studies highlight the interaction between nerve growth and skeletal muscle cells in skeletal muscle regeneration.
Blood vessels supply nutrients and oxygen to muscle tissues. The diffusion distance of oxygen and nutrients from microvessels to cells is only limited to 150-200 µm [91]. Even though immediate hypoxia condition triggers endothelial cell proliferation [92], muscle tissues cannot survive without blood supply in the long term. Endothelial cell (EC)s located on the interior surface of blood vessels are involved in vascular biology. EC transplantation in myocardial injury stimulates angiogenesis, which increases regional perfusion [93]. Vascular endothelial growth factor (VEGF) is a primary regulator of angiogenesis. VEGF-derived angiogenesis promotes skeletal muscle regeneration. VEGF overexpression via virus gene transfer shows an increase in forelimb strength and a decrease in necrotic fibers area in mdx mice [94]. The combined delivery of VEGF and IGF-1 led to angiogenesis, innervation, and SC activation, resulting in functional recovery [95]. In previous research in our lab, transplantation of mesenchymal stem cells into acellular ECM increases blood vessel density, resulting in functional recovery.
CONCLUSIONS
Skeletal muscles can heal in general injuries, but are not capable of regenerating across a large muscle-deficient gap. The repercussions of this severe injury induce subsequent functional deficits due to loss of nerves, vessels, and muscle mass. Moreover, people who sustain VML injury suffer from emotional distress by physical abnormalities and aesthetic deficits. The processes involved in physical and psychological rehabilitation require high medical cost burdens. Current clinical treatments for VML include the transplantation of biocompatible matrix or autologous acellular ECM. However, the transplantation of the scaffolds only has not yet achieved complete functional and morphological recovery. In contrast, SC transplantation for skeletal muscle regeneration is a well-studied medical procedure. Delivery of SCs through single myofiber has shown functional recovery and hypertrophy. RT is a well-known treatment that triggers hypertrophy and increase muscle mass, but the potential interaction between resistance training and muscle cell activation has not been studied yet. Therefore, future studies to identify novel and effective therapeutic treatments of VML injury with myofiber transplantation and/or RT are needed.


