


서론
지구력 능력을 결정짓는 주요 요인 중의 하나는 지방 산화 (fat oxidation)의 증가이다. 증가된 지방 산화는 운동 중 골격근과 간의 글리코겐의 사용을 감소시켜 글리코겐의 고갈의 지연시켜 준다 (glycogen sparing). 여분의 글리코겐은 운동의 후반부에 사용되거나 혈장 글루코스 수준을 유지시켜 더 오랜 시간 운동하는데 도움이 될 수 있다[1]. 골격근의 미토콘드리아에서 베타-산화 (β-oxidation)에 이은 세포 호흡 (cellular respiration)은 지구력 운동 중에 근육 에너지를 위해 적절한 ATP를 생산하는데 필수적이다[2]. 따라서 지방 산화를 증가시키고 탄수화물 소비를 감소시켜줄 수 있는 에너지 대사 조절 능력은 장시간 운동할 때 지구력을 향상시켜 줄 수 있다.
아세트산 (acetic acid, CH3 COOH)은 식이 탄수화물 (eg., cellulose, starch)로부터 장내 혐기성 박테리아 발효 (anaerobic bacterial fermentation)에 의해 생산되는 주요한 단쇄 지방산 (short-chain fatty acid)이다[3]. 장내에서 생성된 아세트산은 혈관으로 흡수되어 아세테이트 (ace-tate)로 전환되어 중성화된다. 젖소와 같은 경우는 대부분의 아세트산은 우유의 지방산 합성에 사용되지만, 사람이나 설치류의 경우 주로 간세포 (hepatocytes)에서 구연산회로 (tricarboxylic acid cycle)를 통해 에너지를 생산하는데 사용된다[4]. 아세테이트는 대부분의 포유동물 조직의 세포 미토콘드리아와 세포질 모두에 존재하는 아세테이트 티오 키나아제 (acetate thiokinase=아세틸-CoA 합성 효소, acetyl-CoA syn-thetase) 효소에 의해 아세틸-CoA로 빠르게 활성화된다[5]. 아세테이트로부터 생성된 아세틸-CoA는 우선적으로 구연산 (citrate) 합성에 사용될 수 있다[6]. 또한 아세트산 투여가 설치류 실험에서 운동 중 지방 산화를 증가시키며 간과 근육의 글리코겐의 축적을 촉진시키며[7], 에너지 소비를 증가시키고 체지방량을 억제시킨다고 보고되었다[8]. 아세트산을 마우스에 투여하였을 때, 간의 중성지방 (triglyceride, TG)을 낮춰주었고 지방 산화와 관련된 유전자들의 발현을 증가시켰다[9]. 또한 당뇨병 동물 모델인 KK-A (y) 마우스에서 아세트산은 간의 AMP-acti-vated protein kinase α (AMPKα)를 활성화시키고 고혈당증 (hyperglyce-mia)을 감소시켰다[10].
AMPK는 계통학적으로 serine/threonine 단백질 키나아제로서 환경적 또는 영양적 스트레스 요인들에 반응하여 지질 대사를 조절하는 대사적 마스터 스위치 (metabolic master switch)의 역할을 하는 조절 단백질이다[11]. AMPK는 급성 및 장시간 운동 중에 골격근의 지방산 대사에 영향을 미치는 중요한 조절 인자이다. AMPK는 여러 지질 대사 관련 전사 인자 (transcription factors)를 조절함으로써 지질 대사를 조절한다. 예를 들어, PPARα (peroxisome proliferator-activated receptor α, SREBP-1c (sterol regulatory element-binding protein 1c), ChREBP (car-bohydrate responsive element-binding protein) 등이 있으며, 이들 모두 전사 인자들은 지질 대사 효소의 발현을 조절한다[12]. 활성화된 AMPK 는 근육 세포로 글루코스 유입 (glucose transport)과 지방 산화 (fatty acid oxidation)의 증가를 자극시키고 에너지 균형을 회복시키기 위해 단백질 합성, 지방 합성, 글리코겐 합성과 같은 ATP를 소비하는 과정 (ATP-consuming process)을 억제시키는 것으로 알려져 있다[13].
아세트산은 또한 근육과 지방 조직에서 AMPKα의 활성화를 통해 지방 산화를 증가시키는 것으로 보인다. 당뇨병 모델인 OLETF 동물에게 아세트산을 투여하였을 때 지방 조직에서 지방 생성 (lipogenesis)을억제시키고 지방 축적을 감소시켰으며 근육 조직으로의 글루코스 유입의 증가와 AMPK α의 활성화에 의해 지방 산화의 증가가 나타났다[14]. 랫 골격근 배양세포 모델인 L6 세포에서 아세트산은 AMPK α의 활성화와 지방 산화와 관련된 유전자들의 발현을 증가시켰고 지방산의 세포 내 유입을 유의하게 증가시켰다[15]. 다른 연구에서 아세트산 투여가 운동 중 골격근의 산화적 인산화 (oxidative phosphorylation)를 증가시키는 것은 AMPK 신호 시스템을 활성화시킴으로 가능하다는 것을 보여주었다[16]. 또한 이 연구에서 아세트산이 근섬유 유형 전환 (muscle fiber type reprogramming)과 운동 중 골격근의 에너지 유형의 전환을 통해 즉, 글루코스 사용보다는 지방산의 소비를 증가시켜 지구력을 향상시킬 수 있다고 제안되었다. 따라서 아세트산은 여러 조직에서 에너지 대사의 조절에 중요한 역할을 하는 것은 분명한 것 같다. 하지만 골격근에서 아세트산에 의한 정확한 대사적 조절 기전이나 골격근의 생리학적 기능, 특히 지구력 향상에 미치는 영향에 대한 정보는 많지 않다. 따라서 본 연구의 목적은 마우스 모델을 통해 아세트산이 지구력 향상에 미치는 효과를 조사하고 지구력과 관련된 생리학적 변인들과 기전들을 조사하여 아세트산이 운동수행능력 제고 보조물로서의 가능성을 찾고자 하였다.
연구방법
1 실험동물
본 연구에 사용된 실험동물은 10주령의 ICR 마우스 수컷 48마리 (샘타코)를 구입하여 1주간의 적응기를 마친 후, 각각 그룹 당 8마리씩 무작위로 임의 배정하여 실험에 사용하였다. 사육실의 온도는 22°C, 습도는 약 50%, 명암은 12시간 주기로 조절하였다. 동물들은 케이지당 4마리씩 사육되었고 사료와 물은 충분히 공급되었다. 실험동물 취급법에 따라 실험하였으며, S대학교 동물실험윤리위원회의 승인을 받아 실시하였다. 초기 동물 그룹은 다음과 같기 구분하였다: Sedentary+control (Sed+Con, n=8); sedentary+sodium acetate (Sed+NaAc, n=8); exercise+ control (Exe+Con, n=8); exercise+sodium acetate (Exe+NaAc, n=8).
2 실험방법
1) Na-Acetate 처치
통제 그룹인 Sed+Con 그룹은 운동을 하지 않고 일반 사료 (chow food)를 먹었으며 Sed+NaAc 그룹은 운동을 하지 않고 매일 5% (w/w, weight of NaAc/weight of chow food)의 NaAc가 첨가된 사료를 먹었다[17]. Exe+Con 그룹은 일반 사료를 먹으며 트레드밀 위에서 1주일에 4일 운동하였고 Exe+NaAc 그룹에 매일 5%의 NaAc가 첨가된 사료를 먹으며 트레드밀 위에서 1주일에 4일 운동하였다. 운동 훈련과 아세트산의 투여는 8주 동안 처치되었다.
2) 지구성 운동훈련 프로그램
지구성 운동훈련 프로그램은 선행연구에서 실시되었던 방법을 활용하여 실시하였다[18]. 8주 실험 기간 동안 동물들은 전동 트레드밀 위에서 1주일에 4일 (월, 화, 목, 금) 훈련을 받았다. 훈련은 10° 경사도에서 10 m/min 속도에서 5 min 동안 실시하고 나서 매 1 min 마다 1 m/min의 속도를 나머지 15 min 동안 증가시켰다. 마지막 20분에서 동물들은 25 m/min의 속도로 달렸다. 달리기를 장려시키기 위해 트레드밀 뒷부분의 전기 충격 grid를 활용하였다.
3) 조직 채취
모든 그룹의 동물들은 CO2 흡입 마취 후 골격근 (전경골근, 비복근 외)과 내장 지방 (visceral fat)을 채취했고, 채취된 조직들은 분석 전까지 −80°C에서 보관하였다. 내장 지방은 epididymal 부위의 지방조직을 오른쪽과 왼쪽 2곳에서 절취하여 무게를 합산하여 개체별 무게 (g/mouse)의 평균으로 나타내었다.
4) 체중과 음식 섭취량
체중은 1주일에 한 차례 측정하여 각 그룹의 평균값을 구하였다. 음식은 1주일에 두 차례 주었으며 음식을 주기 전에 매번 무게를 기록하고 다음 번 음식을 주기 전에 남아있는 음식 무게를 측정하여 음식 섭취량을 측정하였다. 음식 섭취량은 1주일에 두 차례 측정하여 동물 한 마리당 그리고 매일 음식섭취량 (daily food consumption)을 산출하였다.
5) 혈액 분석
동물을 overnight 금식 후, 희생시키기 전 각 동물마다 혈액을 꼬리 정맥에서 채취하여 Accutrend Plus (Roche Diagnostics, Switzerland)를 사용하여 혈액 글루코스 (glucose, mg/dL), 중성지방 (triglycerides, mg/dL), 총 콜레스테롤 (total cholesterol, mg/dL), 젖산염 (lactate, mmol/L) 농도를 측정하였다.
6) 지구력 평가
지구력 평가는 2주에 한 번씩 실시하였다. 지구력 달리기는 10°의 경사에서 처음 5 min 동안 10 m/min의 속도에서 시작하여 스피드가 점차적으로 증가되어 속도는 매 1 min 마다 1 m/min씩 증가시켜 15 min 동안 지속되었다. 동물들은 20 min이 지나고 나서 25 m/min의 속도를 유지하며 탈진될 때까지 달리기 운동을 하였다. 동물의 탈진은 실험자가 브러쉬로 자극하여도 동물들이 20초 동안 전기충격 grid (1.5 mA, 150 V)에서 벗어나지 못하고 머물러 있는 경우로 설정하였다. 탈진된 동물들은 트레드밀에서 제거되어 전체 달리기 시간이 기록되었고 나중에 달리기 거리가 산출되었다.
7) 근 기능 테스트
Kondziella’ s inverted screen test로 불리는 이 검사는 사지 매달리기 검사 (four limb-hanging test)로 설치류 동물의 근력 (muscle strength), 협응 능력 (coordination), 피로에 대한 내성 (tolerance to fatigue)을 측정하는 근 기능 테스트 (muscle function test)로 마지막 8주에 한 차례 실시되었다. 이 테스트는 와이어 그리드 시스템을 사용하여 체중에 반대하는 지속적인 사지 장력을 나타내는 마우스의 능력을 비 침습적으로 측정한다. 동물들은 2분 간격 (휴식)으로 5차례의 매달리기 검사를 받았고 제일 짧은 시간과 제일 긴 시간을 제외한 3차례의 매달리기 평균 시간 (min)으로 근력이 평가되었다.
8) Western blotting 분석
마우스 골격근에의 단백질 측정은 선행연구에서 설명된 것처럼 Bio-Rad사의 Western blot 시스템을 사용하여 전형적인 형태의 전기영동법을 사용하여 특정한 단백질의 양을 분석하였다[19]. 이 실험에서 사용된 Primary 항체: anti-AMPKα, anti-phospho-AMPKα Thr172, an-ti-FAS (Cell Signaling Technology), anti-CPT1B (ThermoFisher Scientif-ic), (Santa Cruz Biotechnology), anti-β-actin (Sigma-Aldrich).
연구결과
1 아세트산이 몸무게, 음식물 섭취량, 내장지방 무게에 미치는 영향
4그룹으로 분류된 동물의 그룹 간 몸무게의 유의한 변화는 실험이 시작되고 4주까지는 나타나지 않았다. 실험이 시작되고 3주까지는 4 그룹 모두 매주 평균 약 0.8 g이 직선적으로 증가하였다. 4주가 되어 Sed+Con 그룹과 Exe+NaAc 그룹 간의 몸무게의 유의한 차이가 나타났고 (p <.05), 두 그룹 간의 몸무게의 차이는 8주가 되어 실험이 끝날 때까지 유지되었다. 6-8주 사이에 Exe+NaAc 그룹과 Exe+Con 그룹 간의 몸무게의 유의한 차이가 나타났다 (p <.05, Fig. 1A). 본 연구에서 8주 달리기 운동 훈련은 몸무게의 감소를 시키지 못하였다. Sed+Con vs. Exe+Con 그리고 Sed+NaAc vs. Exe+NaAc, 이 두 그룹 간의 유의한 차이가 나타나지 않았다 (p>.05, Fig. 1A).
Fig. 1
Fig. 1Effects of 8-week acetic acid feeding and running exercise training on body weight (A) and food consumption (B). Values are means±S.E. ∗ p<.05 vs. Sed+Con, # p<.05 vs. Sed+NaAc.
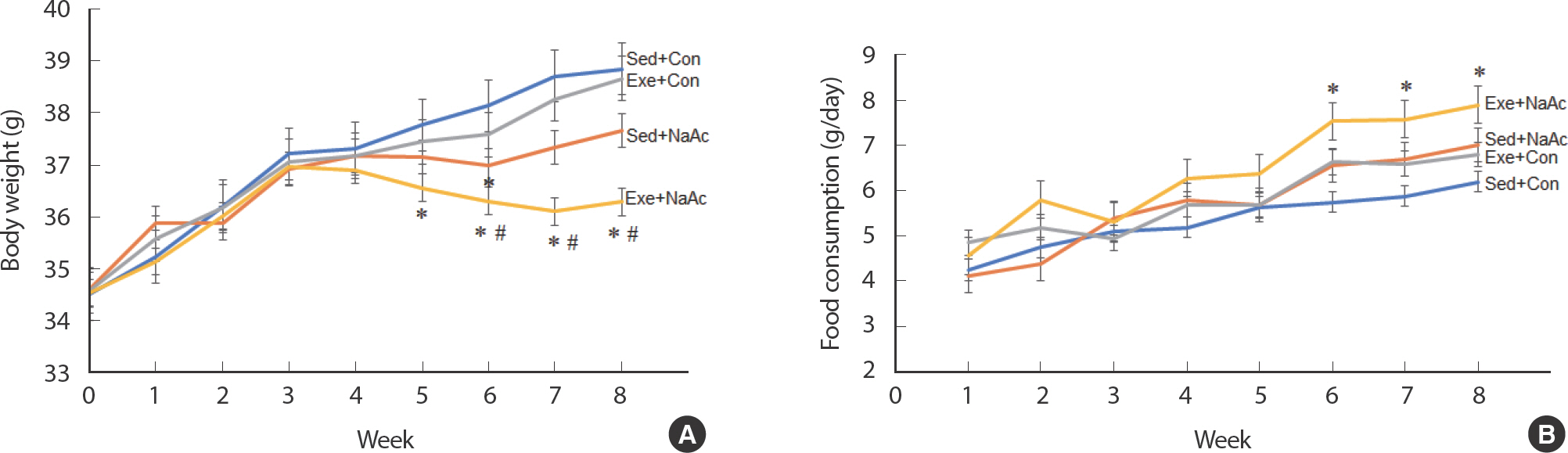
일일 음식 섭취량은 첫 주에 동물 마리 당 평균 약 4.4 g으로 그룹 간의 차이가 없었고 6주째까지는 일일 음식 섭취량의 차이는 나타나지않았다. 일일 음식섭취량은 6-8주 사이에 Exe+NaAc 그룹이 Sed+Con 그룹에 비해 유의하게 증가하였다 (p <.05, Fig. 1B). 6-8주째에 Exe+ NaAc 그룹은 Sed+Con 그룹에 비해, 각각 평균 약 1.8 g 을 매일 더 섭취하였다 (p <.05, Fig. 1B). 몸무게와 일일 음식섭취량의 결과로 보아 6-8주 사이에 Exe+NaAc 그룹은 음식을 더 많이 섭취하면서도 Sed+ Con 그룹에 비해 몸무게의 감소가 유의하게 나타났다.
Sed+NaAc, Exe+Con, Exe+NaAc 그룹들은 Sed+Con에 비해 8주간의 처치에 의해 epididymal fat의 무게가 각각 약 53%, 39%, 59%씩 유의하게 감소하였다 (Table 1). 또한 Exe+NaAc 그룹은 Exe+Con 그룹 보다 epididymal fat의 무게가 추가적으로 감소하였다 (p <.05). Sed+NaAc 와 Exe+NaAc 두 그룹 간의 epididymal fat 무게의 유의한 차이는 없었다 (p>.05).
Table 1
Wet skeletal muscle weight, weight-adjusted muscle weight, epididymal fat weight, and blood parameters
| Sed+Con | Sed+NaAc | Exe+Con | Exe+NaAc | |
|---|---|---|---|---|
| Final body weight (g) | 38.9±0.66 | 37.7±0.56 | 38.7±0.74 | 36.3±0.57∗δ |
| TA wet weight (mg) | 45.6±1.09 | 45.3±0.57 | 45.8±1.79 | 46.0±1.3 |
| TA/BW (mg/g) | 1.18±0.06 | 1.20±0.04 | 1.19±0.07 | 1.27±0.06 |
| Sol wet weight (mg) | 14.7±1.06 | 14.6±0.08 | 14.7±0.67 | 14.6±0.43 |
| Sol/BW (mg/g) | 0.38±0.06 | 0.39±0.05 | 0.38±0.07 | 0.40±0.06 |
| Epididymal fat (g) | 0.99±0.09 | 0.47±0.04∗ | 0.60±0.05∗ | 0.41±0.06∗δ |
| Glucose (mg/dL) | 176.9±12.7 | 187.0±7.6 | 171.1±5.6 | 131.4±6.2∗#δ |
| Lactate (mmol/L) | 3.9±0.2 | 4.1±0.1 | 3.9±0.1 | 3.7±0.2 |
| Triglyceride (mg/dL) | 132.7±5.7 | 101.6±5.3 | 128.1±31.5 | 122.5±10.9 |
| Cholesterol (mg/dL) | 163.9±6.2 | 161.6±3.7 | 167.4±7.7 | 164.9±3.3 |
3 아세트산이 지구력 달리기 시간에 미치는 영향 검사
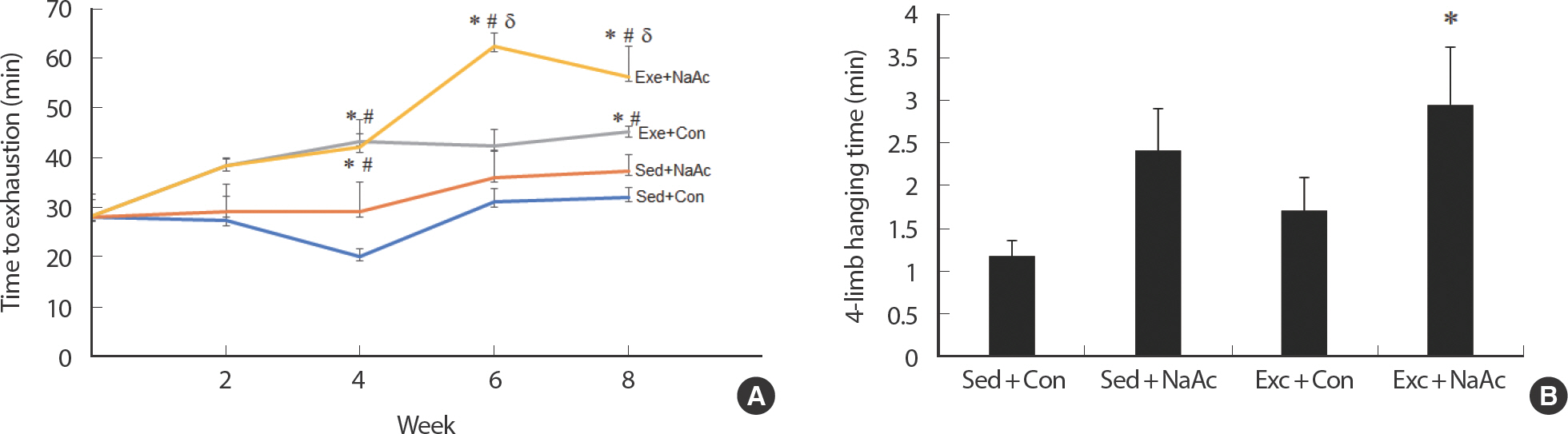
4주째에 Exe+Con 그룹 (−43분)과 Exe+NaAc 그룹 (−42분)의 탈진 시까지 달리기 시간은 Sed+Con 그룹 (−20분)과 Sed+NaAc 그룹 (−29분)에 비해 유의하게 증가하기 시작하였다 (p <.05, Fig. 2A). 6주와 8주 테스트에서 Exe+Con 그룹의 달리기 시간이 Sed+Con과 Sed+NaAc 그룹에 비해 유의하게 증가되었고, Exe+NaAc 그룹은 Exe+Con 그룹에 비해 최대 달리기 시간이 각각 약 110%와 약 70% 증가되었다 (p <.05, Fig. 2A). Sed+NaAc와 Sed+Con 그룹 간의 최대 달리기 시간은 8주 동안 4차례의 테스트에서 한 번도 유의한 차이가 나타나지 않았다 (p>.05).
Fig. 2
Fig. 2Endurance capacity was evaluated by total running time before exhaustion of mice fed with chow food with or without NaAc, with or without exer-cise training for 8 weeks (A). Values are means±S.E. ∗ p<.05 vs. Sed+Con, # p<.05 vs. Sed+NaAc, δ p<.05 vs. Exe+Con. Comparison of four-limb hanging time following 8-week dietary NaAc feeding and treadmill running exercise (B). Each bar represents the means±SE for muscles from 6 mice. (n=8) ∗ p<.05 vs. Sed±Con.
5 아세트산이 지방대사 관련 단백질 효소에 미치는 영향
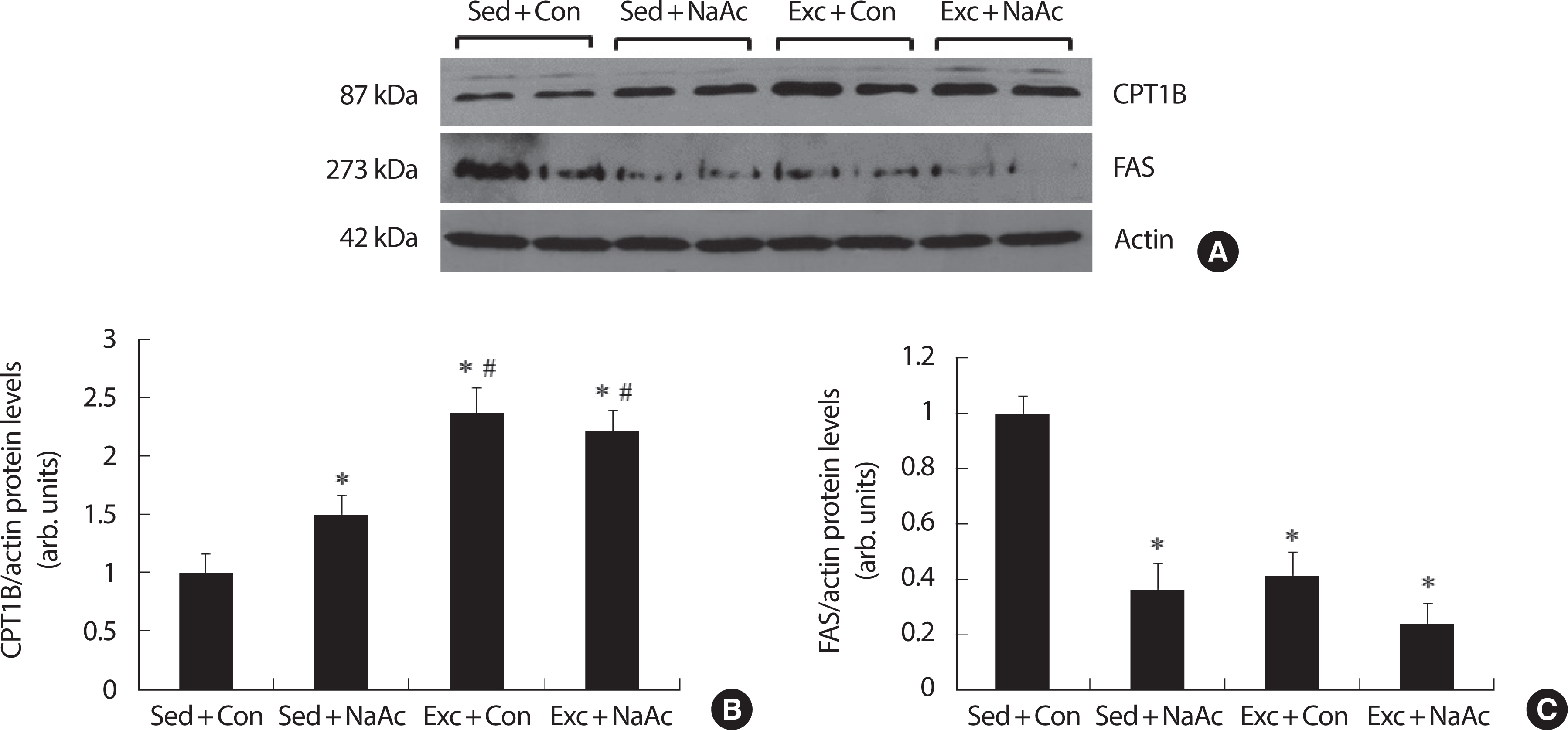
지방산화에 관여하는 단백질 효소 (carnitine palmitoyltransferase 1B, CPT1B)와 지방합성에 관여하는 단백질 효소 (fatty acid synthase, FAS)의 변화를 Western blot 방법을 이용하여 분석한 결과, Sed+Con 그룹에 비해 나머지 모든 그룹에서 CPT1B 단백질량이 유의하게 증가되었다 (p <.05, Fig. 3A, B). 8주 아세트산 투여 (Sed+NaAc)는 CPT1B 단백질 수준을 약 50% 증가시켰고, 8주 지구성 달리기 운동 (Exe+Con)은 CPT1B 를 약 135% 증가시켰으며 8주 아세트산과 운동 훈련 처치는 CPT1B 단백질 수준을 약 120% 증가시켰다. 또한 Exe+Con 그룹과 Exe+NaAc 그룹 은 Sed+NaAc 그룹보다 CPT1B 단백질 수준이 유의하게 증가되었다 (p <.05). 반면 지방산 생합성에 관여하는 효소인 FAS 단백질 수준은 8주 아세트산 투여와 지구성 운동 훈련에 의해 CPT1B 단백질과 반대로 통계적으로 유의하게 감소하였다 (p<.05, Fig. 3A, C). FAS 단백질 수준이 8주 아세트산과 지구성 훈련 처치에 의해 Sed+Con 그룹에 비해 Sed+NaAc 그룹은 약 64%, Exe+Con 그룹은 약 59%, 그리고 Exe+NaAc 그룹은 약 76% 감소되었다 (p<.05).
Fig. 3
Fig. 3Effects of 8-week NaAc feeding and running exercise on CPT1B, fatty acid oxidation marker protein, in skeletal muscle of mice. Representative im-munoblot image of CPT1B, FAS or actin (A). CPT1B/actin (B) was quantitated via densitometry from 8 mice per treatment conditions. FAS/actin (C) was quantitated via densitometry from 8 mice per treatment conditions. Values are means±SE; (n=8) ∗ p<.05 vs. Sed+Col, # p<.05 vs. Sed+NaAc.
6 아세트산이 AMPK 활성화에 미치는 영향
AMPK의 활성화 (thr 172 인산화)는 Sed+Con 그룹에 비해 나머지 3 그룹에서 통계적으로 유의한 증가를 보여주었다 (p <.05, Fig. 4A, B). AMPK의 인산화 수준이 Sed+Con 그룹에 비해 8주 아세트산 투여가 약 4.2배 증가되었고 (Sed+NaAc), 8주 지구성 운동 훈련은 약 6.8배 증가되었으며 (Exe+Con), 8주 아세트산과 지구성 운동 훈련의 복합적 처치는 약 8.2배 증가되었다 (ExevNaAc). 또한 Sed+NaAc 그룹에 비해 Exe+ NaAc 그룹의 AMPK 활성화가 유의하게 증가하였지만 (약 4배, p <.05), Exe+Con 그룹과 Exe+NaAc 그룹의 AMPK 활성화는 통계적으로 유의한 차이가 나타나지 않았다 (p>.05).
Fig. 4
Fig. 4Effects of 8-week 8-week NaAc feeding and running exercise on phosphorylation of AMPK in skeletal muscle of mice. Representative immunoblot images of phosphor-AMPK (thr172), total AMPK or actin (A). Phospho-AMPK (thr172) (B) were quantitated via densitometry from 8 mice per treatment conditions. Each bar represents the means±SE for muscles from 8 mice. ∗ p<.05 vs. Sed+Con, # p<.05 vs. Sed+NaAC.

논의
본 연구는 마우스 동물 모델을 이용하여 8주 아세트산 처치 (5% w/w)가 지구성 달리기 운동 훈련으로 증가된 지구력을 추가적으로 향상시키고 아세트산이 어떠한 기전이나 신호경로를 통하여 지구력 향상을 촉진시키는 가에 대해 조사되었다.
본 연구에서 매주 측정한 동물 몸무게는 아세트산을 섭취했던 그룹 (Sed+NaAc)이 통제그룹 (Sed+Con)과 6주째 한 차례만 차이가 있었을 뿐 나머지 주에서 통계적으로 유의한 차이가 없었다. 아세트산 섭취와 달리기 훈련을 동시에 실시한 동물 그룹은 4주부터 몸무게가 더 이상 증가하지 않고 정체되는 양상을 보인 반면 나머지 그룹들은 꾸준히 증가하는 경향을 보였다. 아세트산과 지구성 훈련 처치는 6주부터 실험이 끝나는 8주째까지 Sed+Con 그룹뿐만 아니라 아세트산을 섭취하지 않으며 지구성 달리기 훈련만 했던 그룹 (Exe+Con) 보다 몸무게를 유의하게 감소시켰다. 따라서 아세트산과 지구성 운동의 복합적 처치는 체중 감소에 유익한 효과를 본 연구에서 보여주었고 이와 같은 체중 감소는 음식 섭취의 감소에 의한 것이 아니라 신체의 에너지 대사의 증가에 의한 것으로 여겨진다. Exe+NaAc 그룹의 동물들은 다른 3그룹들의 동물들보다 오히려 음식을 더 섭취하는 경향을 보였으며 Sed+Con 그룹과는 6주부터 통계적으로 유의한 차이를 보이며 음식 섭취량이 증가되었다. 따라서 Exe+NaAc 그룹은 음식을 더 많이 섭취하면서 몸무게를 더 감소시킬 수 있었다. 이 그룹 (Exe+NaAc)의 동물들은 다른 3그룹들에 비해 더 많은 에너지를 소비하였고, 이 에너지 대사의 증가 는 지구성 운동 훈련과 함께 아세트산을 섭취했을 때에 유익한 효과가 뚜렷이 나타났다. 선행 연구에서 고지방 식이 (high-fat diet)로 비만을 유발시킨 마우스에 아세트산을 6주 투여하였을 때, 음식 섭취량과 골격근 질량의 변화 없이 체지방과 간 지질의 축적이 유의하게 감소되었다[9]. 또한 8주 아세트산을 처치한 마우스 모델에서 통제그룹에 비해 체중이 감소되었고 또한 운동 훈련과 아세트산을 동시에 처치된 그룹이 운동만 처치된 그룹에 비해서도 체중 감량이 나타났다[16].
본 연구에서 동물의 체중 감소는 근육량의 감소가 아닌 내장 지방량의 감소에 의한 것으로 보인다. 동물의 전경골근과 비복근의 무게는 그룹 간의 차이가 없는 것에 비해, 내장지방 (부고환 주위 지방)은 Sed+Con 그룹의 동물에 비해 나머지 모든 그룹들에서 감소하였으며 아세트산과 지구성 훈련의 복합적 처치 그룹 (Exe+NaAc)의 동물들은 지구성 훈련만 했던 그룹 (Exe+Con)에 비해서도 유의하게 내장 지방량을 감소시킬 수 있었다 (Table 1). 따라서 본 연구의 운동 훈련 강도와 아세트산의 농도 조건 하에서 지구성 훈련보다 아세트산이 지방조직에서 지방산의 분해를 촉진시키는 더 강력한 요인인 것으로 여겨진다. 본 연구는 골격근에서 지구성 훈련과 아세트산 투여의 효과를 조사할 목적이었지만 차후 연구에서는 아세트산 투여와 관련해 지방조직에서의 에너지 대사 특히, 지방합성과 지방산화에 관련된 신호경로나 정확한 기전을 조사하는 것은 흥미롭고 관심 있는 연구가 될 것 같다.
본 연구에서 측정한 혈액 지표를 살펴보면 (Table 1), 4그룹들 간의 혈액 젖산, 총콜레스테롤, 혈액 중성지방은 유의한 차이가 나타나지 않았지만, Sed+Con, Sed+NaAc, Exe+Con 그룹들과 비교하여 Exe+NaAc 그룹은 혈당 수치인 혈액 글루코스의 농도가 유의하게 감소되었다. 또한 3그룹들의 평균 혈당 수준이 약 178 mg/dL인 반면 Exe+NaAc 그룹의 혈당 수준은 평균 약 131 mg/dL이었으며 약 −26% (△47 mg/dL) 감소되었다. 따라서 아세트산은 탄수화물 에너지 대사에도 관여하는 것으로 보인다. 선행연구[7]에서 아세트산 투여는 간과 골격근의 글리코겐 저장을 촉진시키는 것으로 밝혀졌다. 한 연구에서 탈진 운동을 시켜 글리코겐을 고갈시킨 후에 아세트산과 글루코스를 동시에 투여했을때 글루코스만 투여한 그룹에 비해 간과 비복근의 글리코겐의 저장량이 유의하게 증가되어 있었다[20]. 따라서 아세트산은 혈중 글루코스 농도를 낮추는 것은 사실인 듯하다.
아세트산은 선행연구와 본 연구의 결과에서 보여주었듯이 지방대사에 관여하는 것 같다. Hattori et al. [8]은 아세트산을 마우스 (C57BL/6J)에 한 차례의 투여한 후 3시간 뒤에 지방산화와 에너지 소비가 증가된 것을 발견하였다. 다른 연구에서 6주 아세트산을 C57BL/6J 마우스에 투여하였을 때, 음식섭취량과 근육량의 변화 없이 체지방량과 간에서 지방 축적이 감소되었고, 이 감소는 간에서 지방산화와 열발생 (thermogenesis)과 관련된 인자들의 발현을 증가시켰으며 이 증가는 PPAR α와 지방산화를 조절하는 AMPKα2의 활성화에 의한 것이라고 제안하였다[9]. 이 두 연구는 아세트산을 투여한 후 간 조직에서 지방대사의 변화를 보았으며 마우스 골격근에서 8주 아세트산의 투여는 지방산화와 관련된 주요한 효소들의 mRNA (ACC2, CPT1β, UCP2/3, HSL) 수준을 통제그룹과 비교하여 유의하게 증가시켰고 이 증가는 골격근의 AMPK 활성화에 의한 것이라고 하였다[16]. 본 연구에서 또한 8주 아세트산의 투여가 골격근의 AMPK 인산화 (thr172)를 증가시켰고 이 AMPK의 활성화가 지방산화와 관련 있는 CPT1B의 발현을 증가시켰을 것으로 여겨진다. 또한 AMPK의 활성화는 근육과 같은 대사성 조직에서 지방의 합성을 억제시키기 때문에 본 연구에서 FAS 효소의 증가는 아세트산에 의한 AMPK의 활성화가 기여한 것으로 여겨진다.
AMPK는 골격근에서 운동에 의해 반응하는 주요한 조절 인자로 알려져 있으며 운동 훈련의 유익한 효과들이 LKB1-AMPK-PGC-1 신호 체계에 의해 이루어지는 것이라 하였다[10]. AMPK 효소는 AMP/ATP ratio의 증가에 의해 활성화되기도 하고 LKB1과 같은 AMPK 상위 키나제 (upstream kinase)에 의한 인산화에 의해 활성화된다. LKB1은 AMPK의 α-subunit의 threonine-172 인산화를 시켜 활성화시킨다[21]. 본 연구를 비롯해 현재 어떠한 연구에서도 아세트산이 정확히 어떻게 AMPK를 활성화시키는 가를 보여주지 못하고 있다. 이러한 기전은 현재 명확히 알려져 있지 않지만 아세트산의 투여는 간과 골격근에서 AMPK를 활성화시켜 지방대사에 관여하는 것으로 여겨진다. 같다.
본 연구에서 8주 지구성 달리기 훈련과 아세트산의 처치는 마우스의 지구성 달리기 능력을 향상시켰다. 8주 달리기 훈련은 통제그룹 (Sed+Con)과 아세트만을 섭취한 그룹 (Sed+NaAc)보다 트레드밀 위에서 지칠 때까지 달릴 수 있는 시간 (최대 달리기 시간)을 유의하게 증가시켰으며 8주 달리기 훈련과 아세트산을 복합적으로 처치하였을 때, 나머지 3 그룹들보다 최대 달리기 시간을 유의하게 증가시켰다. 신체적 훈련을 통한 골격근의 적응은 β-oxidation과 Krebs cycle 효소의 활성을 증가시키며, 근섬유 세포막을 통한 지방산 유입을 증가시킨다. 또한 근육세포에서 carnitine과 carnitine transferase의 활성을 증가시켜 지방산의 유입을 증가시켜서 더 많은 ATP를 공급할 수 있게 한다. 또한훈련에 의해 글루코스 생성과 글리코겐 사용이 감소하게 되고 지방산 산화와 근육 중성지방 사용이 증가하는 것은 미토콘드리아의 크기와 수가 증가하는 것과 일치한다. 즉, 지방 산화에 관련된 효소와 Krebs cycle, 그리고 전자운반연쇄에 의한 대사적 활성화가 증가하게 되면 제한된 글리코겐 사용을 억제시키고, 저장된 지방을 에너지 기질로 사용하게 된다 (glycogen sparing) [22].
아세트산은 간과 골격근에 글리코겐의 저장을 증가시키고 지방산화를 증가시켜주는 것 외에도 아세트산이 운동하는 골격근의 에너지원으로서 중요한 역할을 할 수도 있다. 아세트산 (C2)은 acetyl-CoA로 전환되어 oxaloacetate (C4) 그리고 coenzyme A와 결합하여 citric acid (C6)를 생산할 수 있으며 citric acid는 Krebs cycle을 작용시켜 electron transport system에 전자들을 지속적으로 공급시켜주는 역할을 할 수 있다. 따라서 운동 중 추가적인 아세트산의 섭취는 에너지 능력을 향상시키는데 도움이 될 수 있을 것 같고 특히 지구력 향상에 기여할 수 있을 것 같다. 즉, 아세트산의 투여는 acetyl-CoA를 증가시키고 Krebs cy-cle 그리고 electron transport system을 활성화시켜 근수축에 필요한 더 많은 ATP를 합성하여 본 연구에서 동물의 지구력 검사의 최대 달리기 지속시간을 향상시킬 수 있었을 것으로 여겨진다.
4그룹들 간의 차이가 없을 것으로 예상했던 것과 달리, 8주의 아세트산과 지구성 훈련의 복합적 처치 (Exe+NaAc)는 매달리기 최대 시간을 통제그룹 (Sed+Con)에 비해 유의하게 증가시켰다. 본 연구의 밝혀진 몇 가지 결과의 내용들이 8주 아세트산과 지구성 훈련의 복합적 처치가 피로에 대한 내성을 증가시킬 수 있다는 근거들을 제시해줄 수 있을 것 같다. 특히 지방산화 효소의 증가, 지방합성 효소의 감소, 그리고 근질량의 변화 없이 몸무게 감소는 피로에 대한 내성을 증가시켰을 것으로 보인다. 하지만 현재 선행연구와 본 연구에서 얻어진 결과로는 8주 아세트산과 지구성 훈련의 복합적 처치가 근력과 협응력 향상에 영향을 미쳤다는 것을 정확히 설명하기는 어려울 것 같다.


