Effects of Resistance Exercise and Fermented Soybean Intake on Protein Expression Markers Related to Browning of White Adipose Tissue in Diet-induced Obese Rats
Article information

Abstract
PURPOSE
This study aimed to analyze the combined treatment effect of fermented soybean intake (for additional protein intake) and resistance exercise (for muscle metabolism activation) on the molecular biological response index of white adipose tissue (WAT) browning.
METHODS
The study included twenty 50-week-old male Wistar rats, who were made obese by being fed a high-fat diet for 8 weeks and then were randomly divided into four groups, control (Con), fermented soybean intake (Soy), resistance exercise (RT), and fermented soybean intake and resistance exercise combination (Soy+RT). After 12 weeks of Soy and RT treatment, body composition, and protein expression patterns such as peroxisome proliferator-activated receptor γ coactivator 1-α (PGC-1α), fibronectin type III domain-containing protein 5 (FNDC5), interleukin-6 (IL-6), and uncoupling protein-1 (UCP1) were compared in extensor digitorum longus and retroperitoneal fat.
RESULTS
The obese rats demonstrated a reduction in body weight and body fat mass, as well as maintenance of lean mass. Interestingly, the groups treated with Soy, RT, and Soy+RT displayed enhanced protein expression of PGC-1α, FNDC5, UCP1, and IL-6 in the extensor digitorum longus and retroperitoneal fat tissue than Con. In particular, the Soy+RT group displayed the most remarkable activation of the aforementioned protein levels in the muscle and adipose tissues.
CONCLUSION
This study concluded that resistance exercise and fermented soybean intake can partly contribute to the browning of WAT.
INTRODUCTION
Owing to changes in eating habits and an increase in sedentary life-styles, there has been an increase in the population of obese individuals over the recent decades. Moreover, obesity-related metabolic disorders are now considered a global disease [1]. Considering this, there is now more emphasis on the importance of diet control and exercise programs for effective obesity prevention and treatment [2]. Dietary control through systematically structured nutritional intake and regular execution of an exercise program not only increase energy consumption, but also greatly help increase lean mass [3]. In particular, combined intervention using dietary control through nutritional intake changes and exercise programs shows enhanced effects, along with weight loss, than use of either alone.
White adipose tissue (WAT) browning, which has been reported to be mainly induced by exposure to low temperatures [4] and exercise [5,6], can alleviate insulin resistance and atherosclerosis. These two conditions serve as key factors for the widespread prevalence of metabolic syndrome, due to inflammation in the body that occurs during the process of obesity. Molecular and biological response indicators of WAT browning that appear during the treatment of obesity include peroxisome pro-liferator-activated receptor-gamma coactivator-1alpha (PGC-1α) [7], an indicator of mitochondrial function activation; uncoupling protein-1 (UCP1) [8], a major indicator of thermogenesis; and irisin [9] and inter-leukin-6 (IL-6) [2], representative myokines involved in inflammation and activation of metabolic function.
Effective exercise programs for obesity treatment have mainly focused on aerobic exercise; however, considering the risk of sarcopenic obesity, the importance of resistance exercise, which brings about improvement of muscle function and increase in muscle mass, is also emphasized upon [10,11]. In particular, there is a need for high-intensity interval training or resistance training focused on increasing muscle mass and activating muscle metabolism for browning of WAT [12]. This is based on the fact that resistance exercise improves muscle function and myokine secretion according to muscle mass increase, which can help WAT browning [13]. Therefore, it is necessary to actively seek a browning program for WAT through a nutritional approach for diet control and a com-plex method of resistance exercise for muscle metabolism activation. Despite the benefits listed above, there are still very few studies that have analyzed the effects of such a combined intervention. Protein intake during resistance exercise can induce positive changes in body composition, through increased muscle protein synthesis and increased lean mass [14], while a high-protein diet has an anabolic effect on muscle preservation without the stimulation of resistance exercise [15].
The traditionally used fermented soybean intake method is an effective method for additional protein intake [16]. The isoflavones contained in soybean, which is a representative food of Korea, are known to be effective in preventing osteoporosis and cardiovascular disease, as they have anticancer and estrogen-like activity [17]. They have a biological action similar to that of estrogen, and thus, are considered a food that can be consumed in place of estrogen. In particular, fermented soybeans increase the bioavailability of vitamins, minerals, and isoflavones. In addition, their safety has been recognized, owing to which they are being processed and used as a new food [18]. Fermented soybeans contain various physiologically active substances such as microorganisms and en-zymes, which are produced in the fermentation process of Bacillus strains. Moreover, they also contain the nutritional components of the raw materials, soybeans [19], such as essential amino acids, isoflavones (genistein, daidzein, formononetin, etc.), and various vitamins, which help suppress arteriosclerosis, heart disease, diabetes, and osteoporosis [20,21]. Compared to soybeans, fermented soybeans contain 90 times more leucine, an essential amino acid that inhibits muscle breakdown and promotes muscle synthesis, and are suitable as a protein source for the elderly, because they reduce calcium excretion and get digested well [22]. In addition, they serve as a functional food with the potential to affect immune function via lactic acid bacteria [23]. Soy protein from fermented soybeans has been shown to improve low-density cholesterol concentrations and control dyslipidemia caused by obesity [24].
In view of the above, analyzing the molecular biological response index of WAT browning upon combined treatment with fermented soybean intake (for additional protein intake) and resistance exercise (for muscle metabolism activation) may provide the basis for effective treatment of obesity. To achieve the same, this study analyzed the effect of resistance exercise and fermented soybean intake on changes in the protein expression levels of important browning indicators of WAT in high-fat diet-induced obese rats.
METHODS
1. Subjects
Twenty 50-week-old male Wistar rats were purchased from Doo Yeol Biotech Co. (Seoul, Korea) and used in this study. A high-fat diet (45% fat intake; #D12451, Research diets, Inc., Brunswick, NJ, USA) was applied for 8 weeks, to induce obesity, after 1 week of adaptation to the en-vironment. After obesity induction, the rats were randomly divided into four groups, control (Con), fermented soybean intake (Soy), resistance exercise (RT), and fermented soybean intake and resistance exercise combination (Soy+RT) groups, with five rats in each group. A high-fat diet (45% fat intake) was provided during the 8-week obesity induction period [65]. For 12 weeks, the control group (Con) were treated to 24% casein protein of total calories, the fermented soybean group (Soy) to 24% soybean protein, the resistance exercise group (RT) to 24% casein protein+ladder climbing, and fermented soybean intake and resistance exercise combination group (Soy+RT) to 24% soybean protein+ladder climbing. During the 12-week treatment period, body weight and dietary intake volume were measured every alternate day, at 9 am. Dietary intake volume was measured using the difference between the amount of food provided to each cage and the amount remaining after 48 hours. This study was approved by the Animal Experimental Ethics Committee (approval no. BHCC-IACUC-2016-01) of Daegu Technopark Bio-Health Convergence Center.
2. Diet and exercise treatment
The fermented soybeans were produced domestically. For fermentation, a B. subtilis strain isolated from Deanjang (soybean paste) was used as a seed, and the bacteria were cultured for 9 hours, after 1% inoculation in Luria Bertani broth (Duchefa Biochemie, Haarlem, Netherlands; 10% bacto-tryptone, 5% yeast extract, and 10% sodium chloride), at 37°C. The cultured B. subtilis DJI was centrifuged at 9,950×g and 4°C, for 15 minutes, to recover the cells, following which the recovered cells were washed twice with sterile tertiary distilled water, for use. The soybeans were cleaned and washed with water, immersed in three times the amount of water for 20 hours, steamed for 50 minutes in an autoclave, cooled to 40°C, and inoculated with 1% (v/w) of the raw material, after which the cultured seeds were fermented for 11 hours in an incubator (SW-90S, Sang-Woo Scientific Co., Seoul, Korea), at 37°C.
3. Resistance exercise program
Resistance exercise was performed using a vertical ladder made of wood and iron (1.1×0.18 m, grid 2 cm, and inclination of 80 degrees), with a rest area arranged at the top (20×20×20 cm). The exercise method involved attaching a weight to the tail of the rat while it climbed the ladder. It was performed 3 days a week, for 12 weeks. Before the exercise program, there was an adaptation period in which the ladder climbing was performed 3 times a day, without the weight, for 1 week. After the adaptation period, during the actual exercise program execution period, the weight was added based on the weight of the rats. In the first week, 30% of the body weight; in the second week, 50% of the body weight; and in the third week, 75% of the weight were added to the tail. In the fourth week, 100% of the weight was added, and following that, 25% of the weight was added every two weeks. Ten repetitions were carried out for each exercise, with rests at 1-minute intervals.
4. Tissue sampling
After the 12-week treatment period, the animals were allowed a 48-hour period of recovery, post which their tissues were sampled. After an-esthesia performed by means of injection of Zoletil (0.04-0.06 mL/kg) and Rompun (0.12 mL/kg) into the abdominal cavity, the body composition was analyzed using DEXA, and then the extensor digitorum longus and retroperitoneal fat were rapidly extracted. They were rapidly frozen using liquid nitrogen (−180°C), and stored in a refrigerator main-tained at −70°C, until analysis using western blot. The extensor digitorum longus, selected as the tissue of analysis in this study, is located on the outer side of the lower leg and is connected to the four toes, which makes it the most widely mobilized muscle for toe and dorsal move-ments. Therefore, it is expected that the extensor long extensor muscle will be mobilized as an agonist by the rats in the process of climbing a ladder during the resistance exercise. In addition, retroperitoneal fat, which has been selected as the adipose tissue, corresponds to subcutaneous fat, and constitutes the largest range among the adipose tissue re-gions of rats.
5. Dual X-ray absorptiometry
Zoletil (0.04-0.06 mL/kg) and Rompun (0.12 mL/kg) were administered intraperitoneally to anesthetize the rats, following which the lean mass and body fat mass were measured using Dual X-ray absorptiome-try (DEXA, DSC-3000, Aloka, Tokyo, Japan) in a state where it is well fixed so that it does not move.
6. Western blot
Proteins from the extensor digitorum longus and retroperitoneal fat were analyzed using western blot. After homogenizing the tissue, the amount of protein was measured using Bradford protein assay. The proteins were then separated by means of sodium do-decyl sulfate-poly-acrylamide gel electrophoresis (Bio-Rad, Hercules, CA, USA), transferred to a membrane, blocked with skim milk, and incubated with primary antibodies against GAPDH (Santa Cruz Biotechnology, Dallas. TX, USA), FNDC5 (Cell Signaling Technology, Danvers, MA, USA), IL-6 (Bio-Rad, Hercules, CA, USA) and UCP1 (Santa Cruz Biotechnology, Dallas, TX, USA) (1:1,000 dilution), at 4°C, for 12 hours. After incubation, the membranes were washed three times for 10 minutes using TBST (tris-buffered saline+Tween-20), and then treated with secondary antibody diluted in a ratio of 1:5,000 with blocking solution, for 60 to 70 minutes. After washing 3 times for 10 minutes with TBST, the intensity of the final band was quantified.
7. Statistical analysis
All data were analyzed using SPSS version 20.0, and results have been expressed as mean and standard deviation. After 12 weeks of treatment, two-way analysis of variance and Bonferroni post-hoc test were used to determine the statistical significance of differences between groups, according to the factorial analysis of variance design involving fermented soybean intake and resistance exercise. Results were considered statistically significant at p <.05.
RESULTS
1. Dietary intake and body weight
Changes in dietary intake and body weight during the 12-week treatment period are shown in Fig. 1. Dietary intake increased and decreased repeatedly in all the groups for the first 3 weeks, but there was no significant difference among the groups. However, at the end of 12 weeks, the body weights in the other groups were significantly lower (p <.05) than those in the Con group (Fig. 1).
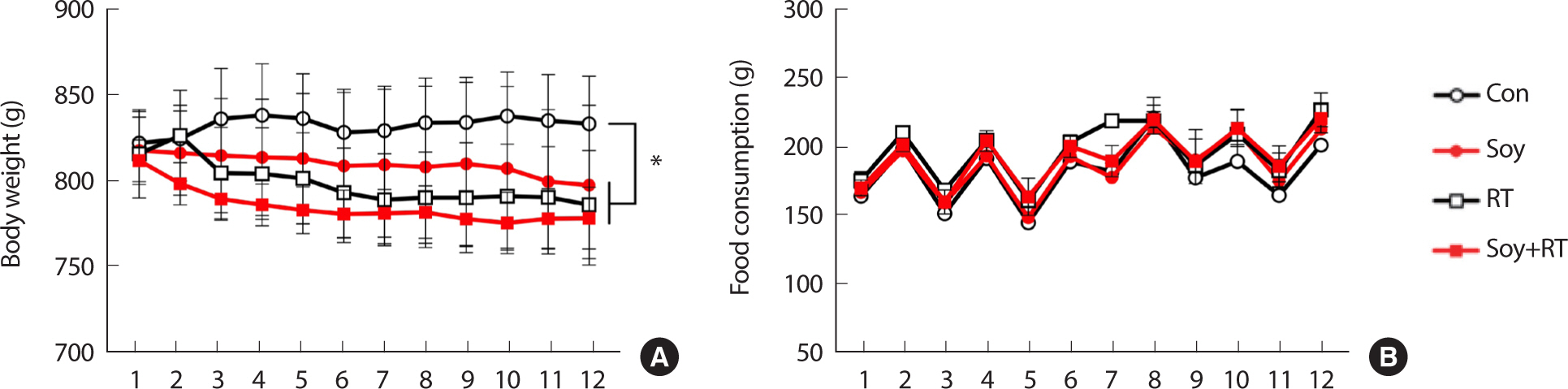
Changes in body weight (A) and dietary intake (B) during the 12-week treatment period (* p<.05 Compared to treatment groups, two-way analysis of variance and Bonferroni post-hoc test, the numbers of each group=5).
2. Body composition
Changes in body composition after 12 weeks of treatment are shown in Fig. 2. Lean mass was significantly (p <.05) higher in the RT and Soy+RT groups than in the Con group. The body fat mass was the highest (p <.05) in the Con group, and the lowest (p <.05) in the Soy+RT group. After 12 weeks of treatment, lean mass to body weight was significantly higher (p <.05) in the other groups than in the Con group, and significantly higher (p <.05) in the Soy+RT group than in the Soy group (Fig. 2).
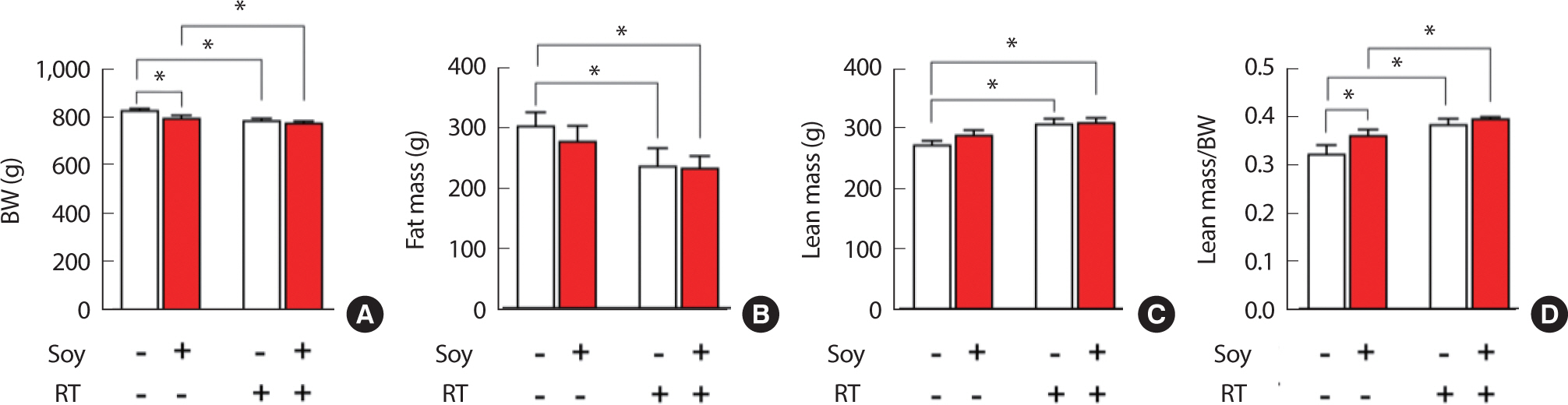
Comparison of body weight (A), fat mass (B), lean mass (C), and lean mass/body weight (D) among the different groups, after 12 weeks of treatment (* p<.05, two-way analysis of variance and Bonferroni post-hoc test, the numbers of each group=5).
3. Protein expression patterns in the extensor digitorum longus muscle
The protein expression patterns in the extensor digitorum longus muscle after 12 weeks of treatment are shown in Fig. 3. The expression of PGC-1α protein was significantly higher (p <.05) in the Soy group than in the other three groups, and significantly higher (p <.05) in the Soy+RT group than in the RT group. However, the same did not differ significantly in the RT and Soy+RT groups, as compared to that in the Con group. The expression patterns of proteins such as FNDC5, UCP1, and IL-6 were significantly (p <.05) higher in the RT and Soy+RT groups than in the Con group. The expression patterns of these protein were also higher in the Soy group than in the Con group, but the differences were not statistically significant (Fig. 3).
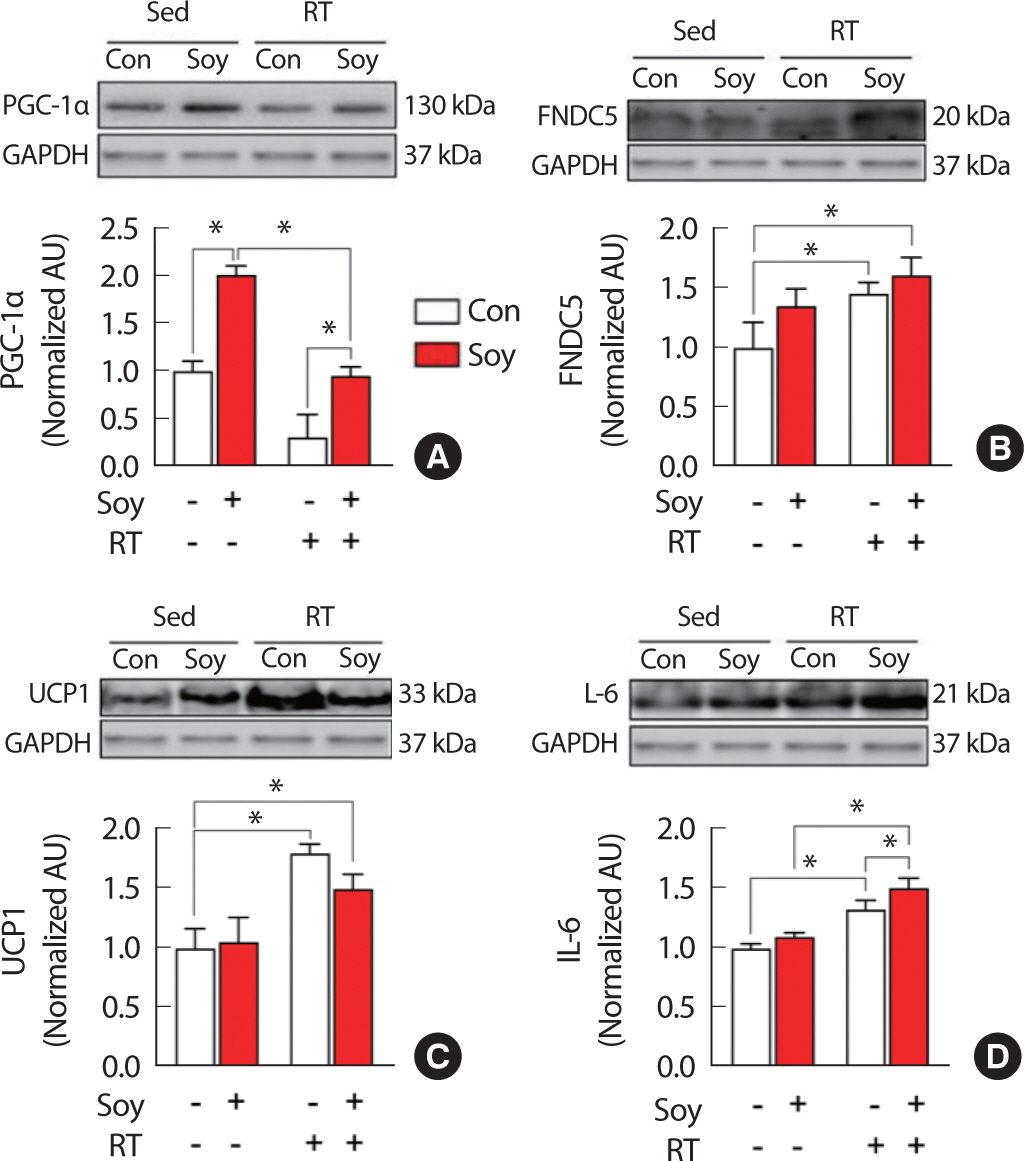
Comparison of protein expression of PGC-1α (A), FNDC5 (B), UCP1 (C), and IL-6 (D) in the extensor digitorum longus muscle, after 12 weeks of treatment.
PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-α; FNDC5, fibronectin type III domain-containing protein 5; UCP1, uncoupling protein-1; IL-6, interleukin-6.
* p<.05, two-way analysis of variance and Bonferroni post-hoc test, the numbers of each group=5.
4. Protein expression in the retroperitoneal fat
The protein expression patterns in the retroperitoneal fat after 12 weeks of treatment are shown in Fig. 4. PGC-1α expression was significantly higher (p <.05) in the Soy group than in the Con group, in the Soy group than in the Soy+RT group, and in the Soy+RT group than in the Con group. However, the same did not differ significantly in the RT and Soy+RT groups, as compared to that in the Con group. FNDC5 expression was significantly higher (p <.05) in the Soy, RT, and Soy+RT groups than in the Con group. UCP1 and IL-6 expression levels were significantly higher (p <.05) in the RT and Soy+RT groups than in the Con group. However, no differences between these protein expression patterns were observed between the Soy and Con groups (Fig. 4).
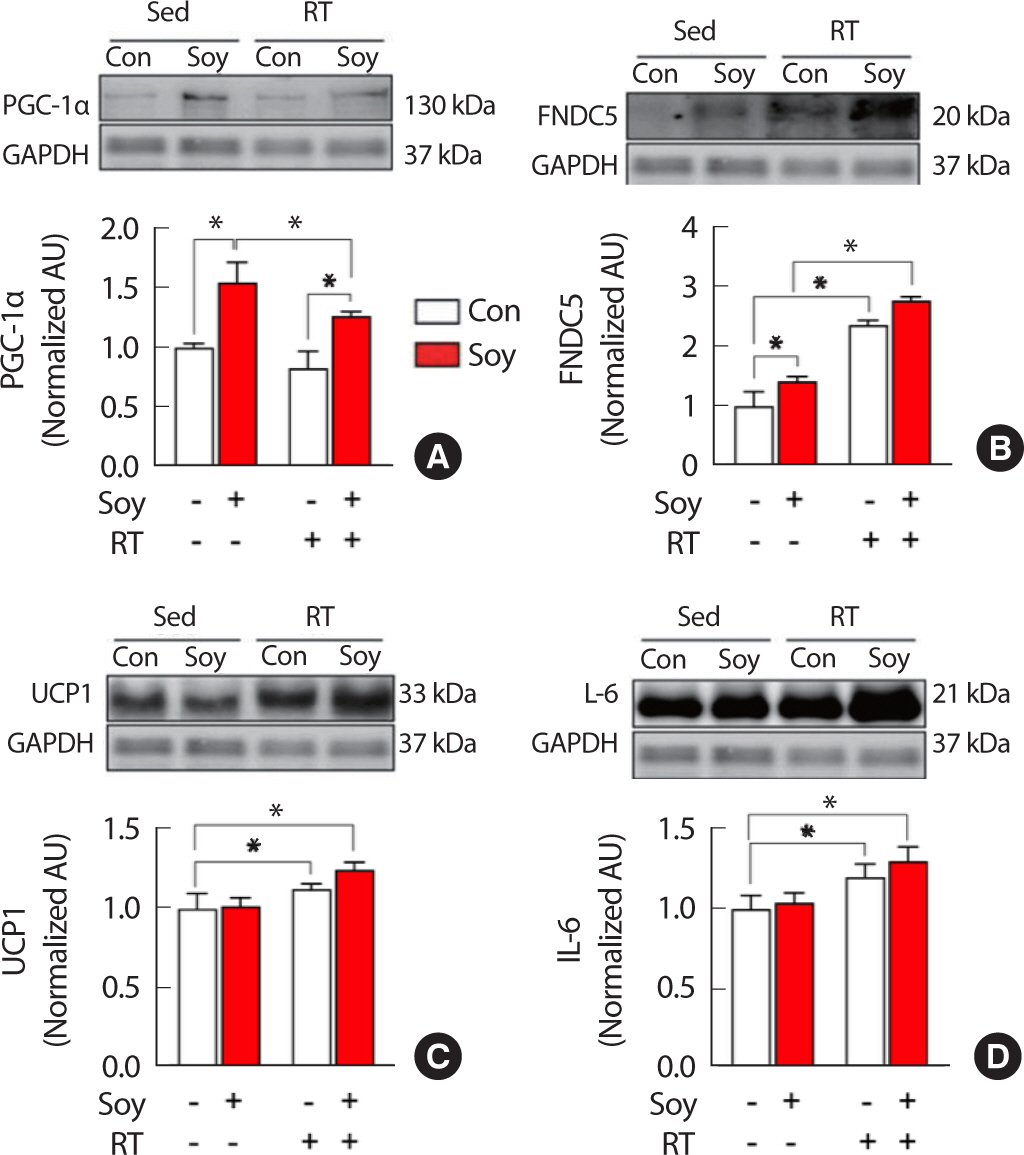
Comparison of protein expression of PGC-1α (A), FNDC5 (B), UCP1 (C), and IL-6 (D) in the retroperitoneal adipose tissue, after 12 weeks of treatment.
PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-α; FNDC5, fibronectin type III domain-containing protein 5; UCP1, uncoupling protein-1; IL-6, interleukin-6.
* p<.05, two-way analysis of variance and Bonferroni post-hoc test, the numbers of each group=5.
DISCUSSION
This study investigated the protein expression patterns of muscle and adipose tissue markers that are considered to affect WAT browning, after resistance exercise and fermented soybean intake in high-fat diet-in-duced obese rats. Resistance exercise and fermented soybean intake caused reductions in body weight and fat mass, as well as a significant increase in lean mass. In particular, the ratio of lean mass to body weight increased significantly in the resistance exercise and fermented soybean intake groups. Therefore, it was confirmed that this combined intervention is very effective in inducing positive changes in body composition. These results suggest that resistance training is an effective method for bringing about an increase in muscle mass [25], and that fermented soybeans can serve a protein source that is more effective in inhibiting muscle breakdown and promoting synthesis than common soybeans [22]. In a study by Ban et al. [26], the group that consumed fermented foods such as gochujang, soybean paste, cheonggukjang, and ssamjang displayed greater weight loss than the high-fat diet group. In particular, it was reported that the group that consumed the food containing soybean paste displayed the most significant weight loss. Moreover, Kim et al. [27] found that the group of rats fed soybean leaf extract for 9 weeks showed more significant weight loss than the control group, in addition to a significant decrease in triglyceride concentration and alleviation of fatty acid synthase protein expression in hepatocytes. Therefore, ingestion of fermented soy protein further activates metabolic activity related to adipose tissue.
PGC-1α, which is expressed in skeletal muscle, affects oxidative stress regulation and weight loss, and is more active during exercise [28]. In addition, PGC-1α plays a key role in the lipolysis of adipose tissue, activation of the thermogenic process in brown adipose tissue, and browning of WAT located in the subcutaneous tissue [29]. In particular, PGC-1α has been considered as an important indicator to stimulate the activation of FNDC5 expression in the process of the browning of WAT [12]. Although the RT and Soy+RT groups did not differ significantly from the Con group in this study, the fermented soybean intake groups were shown to activate PGC-1α protein expression in the extensor digitorum longus. Therefore, consumption of fermented soybeans activates the expression of PGC-1α, which may help in the browning of WAT. However, since the RT and Soy+RT groups did not show a significant difference from the Con group in this study, it can be suggested that the expression pattern of PGC-1α protein in the muscle may differ depending on the exercise type and intake conditions [28,30]. PGC-1α expression in the muscle tissue during endurance exercise is considered a major factor in FNDC5 expression [31]. However, Maak et al. [32] pointed out that the stepwise expression patterns of PGC-1α and FNDC5 proteins during the browning process of WAT may not always be constant for all exercises. Therefore, there is a need for additional analyses to present a clearer con-clusion regarding the stepwise expression patterns of PGC-1α and FNDC5 proteins following resistance exercise and fermented soybean intake.
Mu et al. [12] suggested that browning of WAT through exercise could be an important target in the process of alleviating the risk factors of obesity. Reactive oxygen species, metabolites, the nervous system, exerkines, and lipolysis have been suggested as major mechanisms for the browning of WAT upon exercise intervention. FNDC5 and IL-6, which have been analyzed in this study, are the major exerkines of WAT browning, while UCP1 is a major factor involved in the metabolic process. FNDC5 is a precursor of irisin and acts as a key factor in the thermogenesis and browning of WAT [33]. In both the muscle and adipose tissue, the group subjected to a combination of fermented soybean intake and resistance exercise showed the highest expression levels of FNDC5. Therefore, it was considered that the browning of WAT could be activated through resistance exercise, and a synergistic effect could be further activated through ingestion of fermented soy protein. Previous studies have shown that WAT browning is significantly activated upon admin-istration of FNDC5 to mice [31], while mitochondrial UCP1 expression and WAT browning are significantly activated upon irisin stimulation [34]. Although the results for the response pattern to irisin secretion after exercise have been conflicting, with some studies reporting an increase [35,36], some a decrease [37-41], and some no change [42,43], activation after exercise is considered to be dominant. It is also speculated that the secretion patterns differ according to exercise type, with more studies reporting that resistance exercise leads to higher secretion [44,45], which adequately supports the results of this study. Upon analyzing the effect of regular moderate-intensity exercise after inducing diabetes in rats with ovariectomy, it was found that resistance exercise improves estrogen deficiency, inflammation, and oxidative disorders by increasing serum irisin [46]. Isoflavones in soybeans are also helpful in the prevention and treatment of metabolic diseases caused by obesity, including type-2 diabetes; on similar lines, consumption of fermented soybeans, in combination with resistance exercise, activates the browning of WAT and helps prevent metabolic diseases [47].
UCP1 is a mitochondrial isolation protein that is responsible for the thermogenesis of brown adipose tissue. It increases the conductivity of the inner mitochondrial membrane, thereby allowing the mitochondria to generate heat in brown fat [48]. In particular, activation of UCP1 after exercise induces the expression of important irisin proteins that lead to the browning of WAT, through mitogen-activated protein kinase (P38) and extracellular signal-associated kinase signaling [49]. In this study, the expression level of UCP1 protein in the extensor digitorum longus muscle and retroperitoneal fat was significantly higher (p <.05) in the groups subjected to only resistance exercise or a combination of fermented soybean intake and resistance exercise, than in the control group subjected to only casein intake. Based on this result, it can be deduced that resistance exercise and fermented soybean intake could help the browning of WAT. This could be attributed to the fact that the isoflavone content of fermented soybeans activates UCP1 protein expression in muscle and adipose tissue. In support of our finding, Takashi & Ide [50] reported that the UCP1 mRNA expression in the liver tissue was activated after ingestion of soy protein and isoflavones in rats. Among various approaches, the intake of isoflavone as a type of phytochemical to alleviate obesity in the course of high-fat diet intake can help activate the brown fattening process and lipolysis in WAT [35,47,51]. Many studies have shown that the activation of UCP1 is an important factor in this process [52,53], which is further supported by the finding in the current study that fermented soybean intake could activate the browning of WAT. In particular, the possibility that fermented soybeans have a more significant effect than general soybeans should also be considered [22].
Picoli et al. [13] suggested activation of UCP1 protein expression in the WAT during both endurance exercise and ladder climbing resistance exercise in mice, similar to the activation effect observed in the muscle and adipose tissue after resistance exercise in this study. In addition, Lehnig et al. [54] reported that the UCP1 protein expression pattern dif-fered depending on the area of adipose tissue during resistance exercise in mice, with the level being high in the subcutaneous adipose tissue of the inguinal area. In this study, activation of UCP1 protein expression was observed in retroperitoneal fat, a subcutaneous adipose tissue. Although aerobic exercise has traditionally been recommended for weight control and adipose tissue reduction, resistance exercise has been suggested to increase basal metabolism and decrease visceral fat mass, by increasing muscle mass [55-58]. In addition, resistance exercise has been considered to cause a significant activation of lipolysis in adipose tissue [59]. However, it is unclear whether these results in rats could be extrap-olated to humans, and therefore, there is a need for further studies to as-certain this [60]. The muscle analyzed in this study is EDL, which has a relatively large distribution of type II muscle fibers. Therefore, it can be assumed that the resistance exercise applied in this study had a significant effect, and there will be a need for additional experiments considering the effect of each type of exercise on type I muscle fibers in the future.
IL-6 secreted from the muscles during exercise has positive effects on energy consumption at rest, lipolysis, fat oxidation, and glucose homeostasis [61]. In addition, IL-6 secreted from muscle and adipose tissue is considered a major factor that can help the browning of WAT [62,63]. Li et al. [64] directly administered IL-6 to the hypothalamus of rats, and found that it decreased body weight and visceral fat without affecting changes in food intake, in addition to increasing UCP1 expression in the browning of WAT; this phenomenon, however, was not observed in rats in which the sympathetic nerve acting on brown fat was blocked, thereby suggesting that IL-6 affects UCP1 activation. In the current study, the highest level of IL-6 expression in the extensor digitorum longus muscle tissue was found in the group subjected to a combination of fermented soybean intake and resistance exercise. Therefore, the activation of IL-6 protein expression in muscle and adipose tissue by means of resistance exercise and ingestion of fermented soybeans provides evidence that the same can be treated as an important intervention program for obesity.
In high-fat diet-induced obese rats, resistance exercise and fermented soybean intake helped activate the expression of FNDC5, UCP1, and IL-6 proteins in the muscle and adipose tissue, and thereby possibly caused browning of WAT.
CONCLUSION
Analysis of expression patterns of markers related to the browning of WAT showed that the extensor digitorum longus muscle and retroperitoneal adipose tissue displayed enhanced activation of FNDC5, UCP1, and IL-6 in high-fat diet-induced obese rats treated with both resistance exercise and ingestion of fermented soybeans. Therefore, we deduce that resistance exercise and fermented soybean intake together may facilitate the browning of WAT and help control body weight, while maintaining lean mass, in this animal model.
Notes
The authors declare no conflict of interest.
AUTHOR CONTRIBUTIONS
Conceptualization: K Kim, Y Kim; Methodology: K Kim, J Yoon; In-vestigation: Y Kim; Re-sources: Y Kim, J Yoon; Data curation: Y Kim, G Lee; Writing - original draft preparation: Y Kim; Writing - review and Editing: K Kim; Visualization: G Lee; Supervision: K Kim; Project ad-ministration: K Kim; Funding acquisition: K Kim.