Effects of Lifelong Aerobic Exercise on Ferroptosis-Related Gene Expressions in Kidney of Aged Mice
Article information

Abstract
PURPOSE
This study sought to explore the impact of continuous aerobic exercise throughout life on the expression of ferroptosis-related genes in aging kidneys, elucidating the role of exercise in maintaining kidney health.
METHODS
Male C57BL/6 mice were divided into young control, young exercise, old control, and old exercise groups, with the latter providing access to spontaneous running wheels. Aged mice were observed until 99 weeks, while young mice were followed until 22 weeks. Kidney tissue samples underwent analysis through quantitative polymerase chain reaction, iron assays, histological staining, and pathological damage scoring.
RESULTS
Expression of Nphs1 and Nphs2 exhibited no significant changes after exercise. However, histological analysis revealed that age-related kidney damage, including glomerular mesangial proliferation, was notably offset by lifelong aerobic exercise. The expression of Slc7a11 and Gpx4 remained unaffected by exercise, whereas Nfe2l2 expression decreased significantly in the young exercise groups. Acsl4 expression exhibited no aging - or exercise-induced differences. Additionally, Tfrc expression was lower in the young exercise group than in the young control group but remained consistent between the old and old-exercise groups. Total iron (Fe) increased in aged mice compared to that in the young-exercise group, and the Fe2+/Total Fe ratio was lower in the old-exercise than in the old control group.
CONCLUSIONS
Lifelong aerobic exercise positively impacted kidney health by enhancing antioxidant capacity and mitigating iron accumulation. These findings underscore the significance of exercise in preventing age-related deterioration in kidney function.
INTRODUCTION
The aging process can affect kidney health, including loss of kidney parenchymal volume, glomerular sclerosis, and tubular atrophy, resulting in a decline in kidney function [1]. During the aging process, there is an accumulation of reactive oxygen species and a reduction in antioxidant capacity, which may lead to increased oxidative stress levels and result in kidney damage [2-5]. Furthermore, an increase in iron accumulation is another change that occurs with aging [6]. In the kidneys, it has been noted that iron accumulation increases with aging, and this is known to be highly correlated with lipid peroxidation [7].
Aging-related increases in oxidative stress and iron accumulation may cause ferroptosis [8]. Ferroptosis is a form of regulated cell death that is characterized by iron-dependent accumulation of lipid peroxides within cells [9,10]. This kind of cell death can be inhibited by negative regulators, such as phospholipid peroxidase glutathione peroxidase 4 (GPx4, encoded by Gpx4 gene) and nuclear factor erythroid 2-related factor 2 (Nrf2, encoded by Nfe2l2 gene) [10]. Moreover, it is currently established that ferroptosis is associated with kidney-related pathologies, encompassing con-ditions such as acute kidney injury, chronic kidney disease, and ischemia-reperfusion injury [11,12]. In previous studies, it was reported that the knockout of Gpx4 resulted in acute renal failure in mice [13]. In a diabetic nephropathy mice model, the upregulation of Nrf2 enhances antioxidant capacity and suppresses ferroptosis in the kidney, thereby improving kidney health [14]. Therefore, inhibiting ferroptosis represents a significant target for improving kidney health, and increasing antioxidant capacity is one approach to inhibit ferroptosis.
It is well-established that aerobic exercise represents an effective means to enhance antioxidative capacity [15]. Previous animal studies have shown that aerobic exercise can alleviate ferroptosis by increasing the levels of antioxidative factors including Nrf2 and GPx4, reducing lipid peroxidation, and iron accumulation [16,17]. However, it was reported that 6 weeks of aerobic exercise increased the antioxidant capacity and restored kidney function in 23-month-old rats, while 12 weeks of aerobic exercise was found to be ineffective in enhancing the antioxidant capacity in 22-month-old rats [18,19]. These previous studies show that exercise may suppress ferroptosis in the aging kidney, but it seems that the exercise period is an important factor in suppressing the ferroptosis of the aging kidney.
Previous studies indicate that lifelong aerobic exercise can slow down the aging process and have beneficial effects on skeletal muscles and adipose tissue [20,21]. However, there is limited evidence regarding the influence of lifelong aerobic exercise on kidney health [22]. Consequently, this study seeks to evaluate the impact of lifelong aerobic exercise on ferroptosis-related factors and investigate its impact on kidney health.
METHODS
1. Animals
For this study, 25 6-week-old C57BL/6 (Koatech, Pyeongtaek-si, Gyeonggi-do, Korea) male mice were acquired and raised. Mice were divided into young control (YC, n=7), young exercise (YE, n=6), old control (OC, n=6), and old exercise (OE, n=6) groups. Exercise groups were raised in a cage with a running wheel (Amazon, Changnyeong, Korea) to enable spontaneous aerobic exercise. To be sacrificed on the same day, young mice were raised for 16 weeks starting when the aged mice were 77 weeks old. At the time of sacrifice, the aged mice were 99 weeks old, and the young mice were 22 weeks old (Fig. 1). Mice were raised in an environment maintained at a temperature of 22-24°C and humidity of 50-60% and equipped with a 12-hour automatic light/dark cycle system. Drinking water and food (AIN-93G, Dyets, Bethlehem, PA, USA) were allowed to be consumed freely until the end of the experiment. Every procedure involving animal cares were in accordance with the ethical standards of the Institutional Animal Care and Use and Committee (ap-proval number: PNU-2019-2448).

Study design.
YC, Young control; YE, Young exercise; OC, Old control; OE, Old exercise.
2. Kidney sample collection
To eliminate the acute exercise effects of spontaneous wheel running, running wheels were removed from YE and OE mouse cages 48 hours before sacrifice. After the mice were anesthetized using isoflurane, they were euthanized, and their abdomens were incised to collect kidneys. Kidneys were washed with phosphate-buffered saline, immediately flash frozen using liquid nitrogen, and stored at -80°C until analysis.
3. Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qPCR) was performed to analyze the messenger RNA (mRNA) level of Nphs1 (NPHS1 adhesion molecule, encoding Nephrin), Nphs2 (Nephrosis 2, encoding Podocin), Gpx4, Nfe2l2, Scl7a11 (solute carrier family 7 member 11, encoding Slc7a11), Acsl4 (acyl-CoA synthetase long chain family member 4, encoding Acsl4) and Tfrc (transferrin receptor 1, encoding TfR1). 30 to 50 mg kidney tissues were homogenized by using a TissueLyser II homogenizer (Qiagen, Hilden, Germany) with steel beads and TRIzol reagent (#15596018, Ambion, Carlsbad, CA, USA) in a 1.5 mL tube with round bottom. After homogenization, total RNA was isolated using phenol pre-cipitation [23]. The concentration and purity of total RNA were measured using a spectrophotometer (Nanodrop 2000c, Thermo Fisher Scientific, Waltham, MA, USA). Complementary DNA was synthesized from 1 ug RNA using AccuPower® RT Premix (K-2041, Bioneer, Daejeon, Korea). To analyze gene expression, synthesized complementary DNA, forward and reverse primers, DEPC-DW, and Fast SYBR TM Green Master Mix (4385612, Applied Biosystems, Foster City, CA, USA) were mixed. The mixed samples were distributed in triplicate in a 96-well plate (4346907, Applied Biosystems, Foster City, CA, USA). Actb was used as an endoge-nous control to normalize mRNA expression levels between samples. Amplification and analysis were performed using the StepOnePlus Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). The primer sequences for each gene used in the experiment are shown in Table 1. The relative expression level of mRNA was calculated using the 2-ΔΔC(T) method [24].

Primer sequences of target genes for qPCR analysis
4. Iron assay
To determine the degree of iron accumulation in kidney tissue, an Iron Assay Kit (ab83366, Abcam, Cambridge, MA, USA) was used. 10 mg of kidney tissue was washed of impurities in cold phosphate-buffered saline and homogenized in iron assay buffer. Centrifugation was performed at 16,000 × g for 10 minutes, and the supernatant was transferred to a 1.5 mL tube. To measure Fe2+, 30 µL sample and 75 µL iron assay buffer were added to each well. To measure Total Fe, 30 µL sample, 70 µL iron assay buffer, and 5 µL iron reducer were added to each well. After incubation at 37°C for 30 minutes, 100 µL iron probe was added and incubated for 60 minutes at 37°C with light blocked. Then, the absorbance at 593 nm was measured using a microplate reader (Emax, Molecular Devices, Sunnyvale, CA, USA). All samples were measured for Fe2+ and Total Fe. Results of Total Fe were expressed as absorbance, and absorbance was used to calculate the Fe2+/Total Fe ratio.
5. Hematoxylin & Eosin staining
Hematoxylin and eosin staining (H&E staining) was performed on frozen kidney section. Kidney tissue was embedded in Tissue-Tek OCT compound (Sakura Finetek, Torrance, CA, USA), rapidly frozen in liquid nitrogen, and prepared to a thickness of 8 um using a cryosection ma-chine (Lecia CM1950, Leica Biosystems, Wetzlar, Germany). The sections were rinsed once in 95% ethanol and fixed with 10% formalin. Afterward, the sections were stained with hematoxylin for 30 seconds, rinsed twice in distilled water, and once in 95% ethanol, followed by eosin staining for 15 seconds. Then, after dehydration and clearing in 95% ethanol, 100% ethanol, and xylene, it was covered with a cover slip.
6. Kidney pathological damage score
After H&E staining, the sections were observed under a microscope (Zeiss Axio Observer Z1, Zeiss, Oberkochen, Germany), 10 randomly se-lected locations were photographed at ×200 magnification. Three experts evaluated the degree of kidney damage using the images taken. The evaluation consists of the degree of glomerular proliferation expansion and renal tubulointerstitial score. The detailed evaluation is as follows [25]:
a. Degree of glomerular mesangial expansion: no expansion=0 point; Mild dilatation=1 point; moderate dilatation=2 points; Severe dilatation=3 points.
b. The tubulointerstitial score is assessed by four indicators such as tubular degeneration and necrosis, tubular atrophy, infiltration of interstitial inflammatory cells, and interstitial fibrosis: None=0 point; <25%=1 point; 25-50%=2 points; >50%=3 points.
7. Statistical analysis
Statistical data analysis was performed using GraphPad Prism 9 soft-ware (GraphPad Software Inc., San Diego, CA, USA). One-way ANOVA was used to analyze qPCR results, histological kidney damage scores and Total Fe, and the Tukey method was used for post hoc analysis. Indepen-dent-sample t-test was used to analyze the Fe2+/Total Fe ratio results. The statistical significance level was set at p <.05, and all data were presented as mean and standard deviation.
RESULTS
1. Effects of aging and lifelong aerobic exercise on kidney function-related gene expressions
qPCR was performed to confirm renal filtration function factors. There was no significant difference in the expression of Nphs1 between YC and OC. YE and OE showed no significant difference from the respective control groups (YC, OC) (Fig. 2A). There was no significant difference in the expression of Nphs2 between YC and OC. YE and OE were not significantly different from the respective control groups (YC, OC) (Fig. 2B).
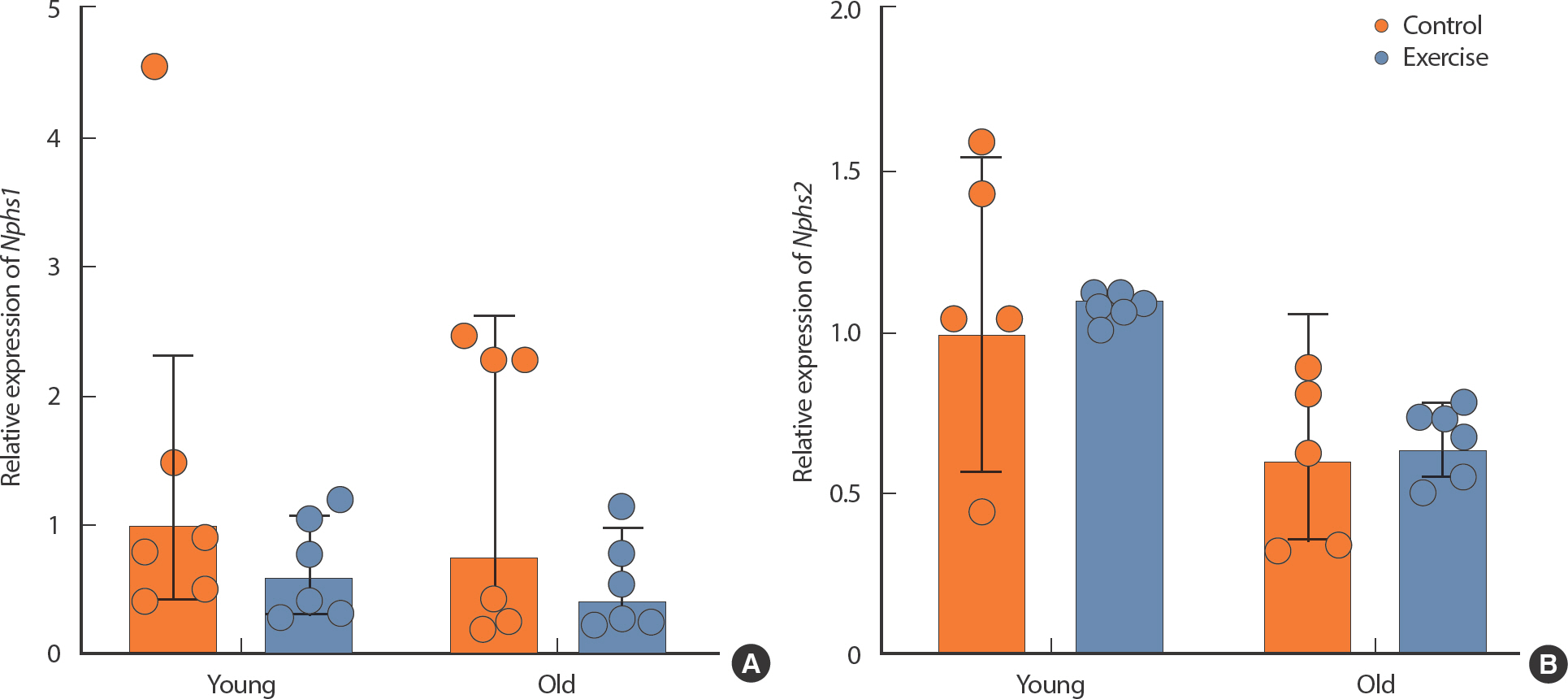
The effect of exercise in kidney function-related mRNA expression in different age in mice kidney. (A) Nphs1, (B) Nphs2. All data are presented as mean±standard deviation (SD).
2. Effects of aging and lifelong aerobic exercise on structure of glomeruli and tubules in kidney
H&E staining was performed to confirm the structure of glomeruli and tubules (Fig. 3A). In terms of glomerular mesangial expansion scores, both OC and OE were significantly higher than YC and YE (p <.05) (Fig. 3B). OE was significantly lower than OC (p <.05) (Fig. 3B). And in the tubulointerstitial score, both OC and OE were significantly higher than YC and YE (p <.05) (Fig. 3C). However, there was no significant difference between OC and OE (Fig. 3C).

Pathological damage score in kidney. (A) Representative images of H&E staining of kidney tissue. (B) Glomerular mesangial proliferation. Normal glomerular (black arrow) and damaged glomerular (yellow arrow) were shown. (C) Renal tubulointerstital score. All data are presented as mean±SD.
** p<.01, vs. control of same age, # p<.05 vs. control of different age.
3. Effects of aging and lifelong aerobic exercise on ferroptosis in kidney
qPCR was performed to confirm ferroptosis factors. The expression of genes related to antioxidants, Slc7a11, Gpx4, and Nfe2l2, was measured. There was no significant difference in the expression of Slc7a11 and Gpx4 regardless of aging and exercise (Fig. 4A, B). However, in the case of Nfe2l2, the expression was significantly lower in OC than in YC, and the expression was significantly lower in YE than in YC (p <.05) (Fig. 4C). Acsl4, a gene related to lipid peroxidation, showed no significant difference according to age and exercise (p <.05) (Fig. 4D). In the case of Tfrc, which is in-volved in the influx of iron into cells, there was no significant difference between OC and OE, but its expression was significantly lower in YE than in YC (p<.05) (Fig. 4E).

The effect of exercise in ferroptosis-related mRNA expression in different age in mice kidney. (A) Slc7a11, (B) Gpx4, (C) Nfe2l2, (D) Acsl4, (E) Tfrc. All data are presented as mean±SD. Slc7a11, solute carrier family 7 member 11; Gpx4, glutathione peroxidase 4; Nfe2l2, NFE2 like bZIP transcription factor 2; Acsl4, acyl-CoA synthetase long chain family member 4; Tfrc, transferrin receptor 1.
** p<.01, vs. control of same age; # p<.05 vs. control of different age.
4. Effects of aging and lifelong exercise on accumulation of Fe2+ in kidney
The Iron Assay Kit was used to determine the degree of iron accumulation in kidney tissue. The level of Total Fe was significant increase in OC and OE compared to YC (p <.05) (Fig. 5A). There was no significant difference between YC and OC in the Fe2+/Total Fe ratio, but a significant decrease was observed in the OE compared to OC (p <.05) (Fig. 5B).
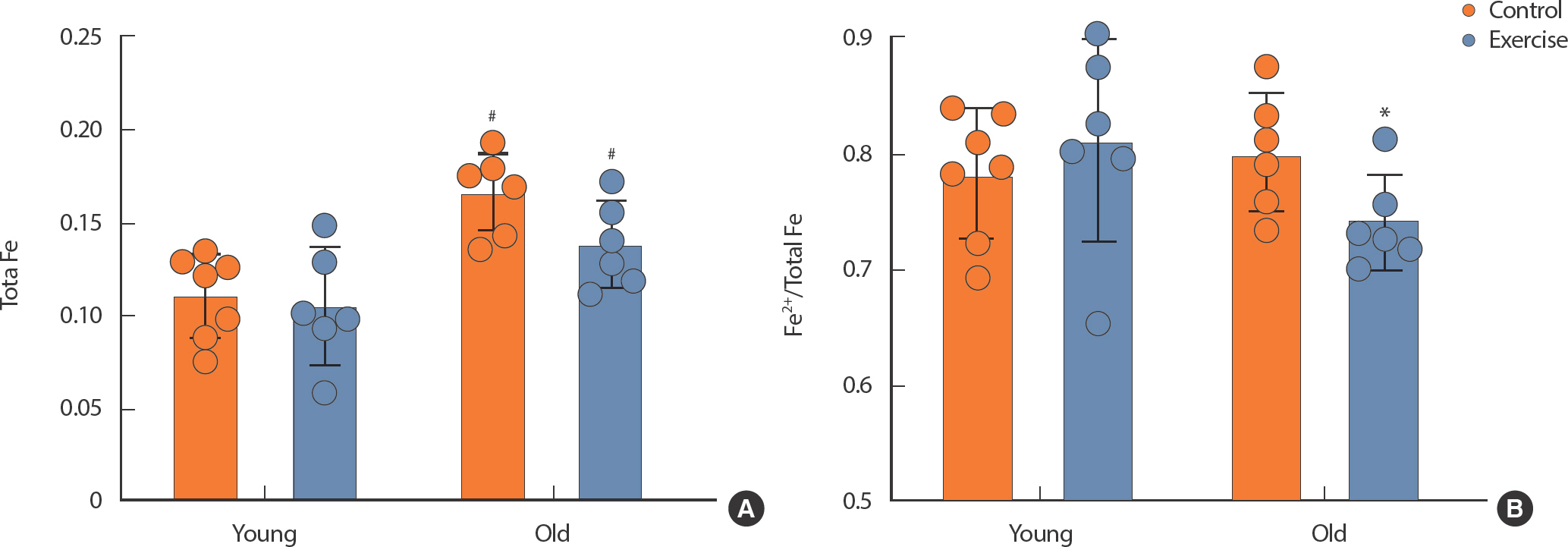
The level of Fe in kidney tissue. (A) Total Fe, (B) Fe2+/Total Fe. All data are presented as mean±SD.
# p<.05 vs. control of different age, * p<.05, vs. control of same age.
DISCUSSION
The kidney is subject to the accumulation of iron and heightened oxidative stress as a result of the aging process, culminating in the onset of ferroptosis, which in turn contributes to kidney dysfunction. Consequently, the amelioration of iron accumulation and the inhibition of ferroptosis within the kidney are pivotal in mitigating aging-related kidney damage. This research sought to evaluate the impact of lifelong aerobic exercise on the modulation of ferroptosis-related factors in aging kidneys.
Histological analysis confirmed the occurrence of kidney glomerular damage and tubular atrophy owing to the aging process. Encouragingly, the aging-related glomerular impairments were significantly decreased through long-term exercise, as illustrated in Fig. 3. Previous studies on rats that underwent lifelong spontaneous wheel running over an 18-month span have also substantiated the positive effects of such exercise on kidney morphological changes [22]. Nevertheless, it is worth noting that the previous research predominantly focused on histological changes in the kidney and did not encompass an evaluation of molecular alterations. In this regard, our current study complements this research by conducting an analysis of gene expression linked to kidney function.
Notably, the expressions of Nphs1 and Nphs2 genes, pivotal in the for-mation of the slit diaphragm and crucial for renal filtration function, dis-played no significant changes attributed to aging or exercise in our study (Fig. 2). Previous research had suggested that aging decreased the expressions of these genes, while aerobic exercise exerted the opposite effect [26-28]. Intriguingly, our study did not identify such positive changes in Nphs1 or Nphs2 expressions related to lifelong spontaneous wheel running. Since Nphs1 and Nphs2 are the biomarkers of kidney damage, it is thought that Nphs1 and Nphs2 do not increase due to there was damage to the kidney even after lifelong exercise [29]. Nevertheless, it was reassuring to observe that the damage to aging kidneys was indeed attenuated by exercise (par-ticularly in the damaged glomerular), as highlighted in Fig. 3.
Furthermore, our study delved into the role of Nfe2l2 (Nrf2), a significant player in intracellular lipid peroxidation and iron regulation [30]. In general, Nfe2l2 (Nrf2) is known to increase with exercise and decrease with aging [30]. In our study, we discovered a substantial reduction in the expression of the antioxidant factor Nfe2l2 due to aging, but this reduction was also observed in YE (Fig. 4C). Similarly, Gpx4, a gene regulated by Nrf2, exhibited a trend mirroring Nfe2l2 (Fig. 4B). The expression of Nfe2l2 is affected by oxidative stress [31]. Immediately after exercise, the expression of Nfe2l2 in skeletal muscle is transiently increased in response to oxidative stress induced during exercise [31]. However, the expression of Nfe2l2 did not show an increase post-exercise after antioxidant pre-treatment [31]. Therefore, it is believed that the decrease in Nfe2l2 expression in YE may be due to a decrease in oxidative stress at rest.
It has been reported that exercise increases Nrf2 protein expression in the kidneys of aged Fischer 344 mice after a short period of 6 weeks of exercise, but many other studies have reported no significant difference [18]. However, it should also be noted that in experiments performed on rats, the beneficial effects of exercise in terms of reducing oxidative stress were evident in 13-month-old rats, but not in 19.5-month-old rats [32]. This fact suggests that differences between mice and rats, as discussed in the 2009 study by George et al. [33] as well as the influence of exercise proto-col cannot be ruled out. In particular, unlike OE, the OC mice used in this study did not exercise at all and had a significantly high mortality rate [21]. Therefore, we cannot rule out the possibility that the difference in Nfe2l2 (Nrf2) expression between OC and OE was underestimated be-cause most of the OC mice with significantly low Nfe2l2 (Nrf2) expression died early and tissues were not obtained. Nevertheless, it is clear that exercise-induced expression of Nrf2 may be impaired due to aging [34]. This could potentially be the primary reason why the expression pattern of Nfe2l2 was different between young and old mice in our study. Our study also shows that the expression of Nfe2l2 in mice acclimated to lifelong voluntary exercise is likely to be offset by aging. In summary, research findings, including those from this study, demonstrate that the expression of Nrf2 is impaired by aging, and this impairment counteracts Nrf2 signaling induced by exercise. While some studies have suggested that exercise-induced Nrf2 signaling can be counteracted by damage, there are other studies that report opposing results. Therefore, it is essen-tial to adopt a multifaceted approach in future research efforts, which in-tegrates a sports nutrition approach to ascertain the optimal frequency, intensity, time, and type of exercise to rescue the Nrf2 pathway that may be compromised by aging.
Antioxidant factor Slc7a11 exhibited an increase with aging, while long-term exercise augmented the expression of this gene (Fig. 4A). Con-versely, the expression of the lipid peroxidation-promoting factor Acsl4 experienced a slight decrease as a result of exercise (Fig. 4D). Notably, Slc7a11 plays a role in diminishing the accumulation of reactive oxygen species and serves as a negative regulator of ferroptosis, whereas Acsl4 can induce ferroptosis by promoting lipid peroxidation of the cell membrane, thereby resulting in cell membrane rupture [10]. Previous research has demonstrated that moderate-intensity treadmill exercise over four weeks elevated brain Slc7a11 protein expression and concurrently low-ered Acsl4 protein expression in a traumatic brain injury model [17]. This study, however, primarily employed healthy mice, which might ex-plain the lack of a substantial effect on lipid peroxidation inhibition due to exercise. Additionally, a study by Wang et al. indicated that lifelong exercise culminated in decreased Slc7a11 protein and Acsl4 gene expression in skeletal muscle [35]. Accordingly, the lifelong spontaneous wheel running regimen in our present study is likely to have enhanced antioxidant capacity by increasing Slc7a11 and decreasing Acsl4.
Moreover, our research discerned a decrease in the expression of the iron regulatory factor Tfrc in YE compared to YC, with no discernible aging-related differences (Fig. 4E). In addition, our analysis of kidney tissue revealed a notable increase in the Total Fe level in aged mice compared to YC. Total Fe has been reported to increase with aging, which may lead to kidney tissue damage [7,36]. Therefore, in this study, the glomerular proliferation expansion was probably associated with increased Total Fe. Compared to OC, the reduction in the Fe2+/Total Fe ratio was shown in OE. It suggests diminished iron accumulation. Tfrc is integral to the regulation of iron influx into cells, and a reduction in Tfrc expression would lead to decreased Fe2+ in kidney tissue [37]. Remarkably, the reduction in Tfrc expression attributed to exercise primarily manifested in the younger mice and was less pronounced in the older age group. In light of this, we believe that spontaneous exercise throughout life played a constructive role in preventing the accumulation of Fe2+ accumulation in aging mice, as evidenced in Fig. 5. Put differently, long-term exercise holds the poten-tial to counteract aging-induced ferroptosis by diminishing Fe2+ levels.
In summary, our research illustrates that although kidney damage is a hallmark of the aging process, lifelong aerobic exercise can effectively prevent aging-related kidney damage by enhancing antioxidant capacity within the kidney and reducing Fe2+ accumulation, thereby preventing the progression of kidney ferroptosis. Subsequent studies should endeav-or to validate the impact of exercise not only at the gene level but also at the protein level.
CONCLUSIONS
This study suggests that lifelong aerobic exercise may help in maintaining kidney health by increasing antioxidant capacity and reducing Fe2+ accumulation, thus contributing to resistance against ferroptosis in aged-kidneys.
Notes
The authors declare that they do not have conflict of interest.
AUTHOR CONTRIBUTIONS
Conceptualization: YY Xiang, KW Baek, JS Kim; Data curation: YY Xiang, KW Baek, JH Won; Formal analysis: YY Xiang, KW Baek; Funding acquisition: KW Baek, JS Kim; Methodology: YY Xiang, KW Baek, JH Won; Project administration: JS Kim; Visualization: YY Xiang; Writing - original draft: YY Xiang, JS Kim; Writing - review & editing: KW Baek, Y Park, JS Kim.