운동훈련과 단백질 섭취에 따른 골격근 단백질 대사: 안정성 동위원소 추적체법을 이용한 연구결과를 중심으로
Review on Exercise Training and Protein Intake in Skeletal Muscle Protein Metabolism
Article information
Trans Abstract
INTRODUCTION
Regulation of skeletal muscle protein mass is implicated not only in exercise performance but in metabolic health. Exercise in combination with nutrition, particularly dietary protein/amino acid intake, are the pragmatic approach that effectively induces muscle anabolic response (i.e., muscle hypertrophy) through regulating protein synthesis and breakdown.
PURPOSE
The purpose of this review was to summarize available data on the effect of exercise intervention and amino acids intake on muscle protein synthesis and breakdown and provide an insight into development of an effective exercise intervention and amino acids supplements, applicable to training practice.
METHODS
In this review, we have reviewed currently available data mainly from stable isotope tracer studies with respect to the effect of exercise intervention and protein or amino acid supplement on muscle protein anabolic response.
CONCLUSIONS
Taken together, exercise alone may not be effective in achieving a positive net muscle protein balance due to the fact that protein breakdown still exceeds protein synthesis until nutrition intake such as protein/amino acids. It appears that muscle anabolic response increases in proportional to the amount of protein intake up to 20 – 35 g depending on quality of protein, age, differences on exercise intensity, duration, and frequency, and individual’s training status.
서 론
골격근은 제지방 조직의 가장 많은 부분을 차지하며, 신체 이동, 자세와 체온을 조절하는 역할을 하고 인슐린에 반응하여 혈당을 흡수(uptake)하는 조직이다. 골격근량은 어린이와 청소년 및 젊은 성인의 ≥ 85%[1,2], 중년과 그 이상 성인들의 50-80%가 유전적인 요인에 의해 결정되는 것으로 추정될 만큼 유전적인 경향이 강하지만[3], 골격근은 매우 역동적(dynamic) 기관으로 크기가 빠르게 증가하거나 감소할 수 있다.
골격근 단백질은 지속적으로 대사회전(turnover), 즉 근단백질 합성(muscle protein synthesis, MPS)과 분해(muscle protein breakdown, MPB)를 일으키며, 단백질합성과 단백질분해의 상대적인 속도에 의해서 단백질 양이 결정된다. 예를 들어, 근단백질량이 변하지 않는다면, 이는 근단백질 합성(MPS)속도와 분해(MPB)속도가 동일하다는 것을 의미하고 대부분의 성인들이 이 상태에 있다고 할 수 있다. 근단백질량의 변화는 근단백질 합성속도와 분해속도가 차이가 있을 때, 즉 합성 속도가 분해속도보다 상대적으로 클 때 근비대가 일어나고 반대일 경우에는 근위축이 일어나게 된다[4,5].
건강한 성인들의 경우, 근단백질의 대사회전율(turnover rate)은 하루에 1-2% 정도로 보고된다[6]. 근단백질 대사회전율은 단백질합성과 분해에 관여하는 신호전달기전의 활성도, 유전인자의 전사 및 단백질 분해 관련 효소 등의 작용이 관여한다[6]. 운동은 이들에 영향을 미치는 주요한 인자로[7], 운동은 근단백질 합성(MPS)을 40-150%까지 증가시킨다[8]. 또한 운동은 테스토스테론, 인슐린, 성장호르몬 및 인슐린 유사 성장인자(insulin-like growth factor, IGF-1)와 같은 근단백질 합성과 관련된 것으로 보고된 호르몬의 변화도 유도하여 근단백질 합성속도에도 영향을 주는 것으로 보고되었다[9]. 그러나 공복상태에서의 운동은 근단백질 분해(MPB)도 증가시키므로 순 단백질균형(NPB)은 여전히 부적상태(분해>합성)에 있게 된다[10]. 아미노산이나 단백질의 섭취를 통한 혈중 아미노산 가용도(availability) 증가도 근단백질 합성(MPS)을 자극하여 순 단백질균형(NPB)을 증가시키지만[7], 저항성운동과 영양소(특히 필수아미노산) 섭취 병행은 보다 효과적으로 단백질 균형을 증가시키므로[11] 저항운동과 아미노산/단백질을 같이 병행하는 것이 바람직하다.
그러나 저항운동에 따른 동화작용 효과는 오랫동안 연구되어 왔으며 근력과 근비대와 관련된 많은 효과들이 입증되었음에도 불구하고 운동과 아미노산/단백질 섭취에 따른 근단백질 합성과 분해에 대한 급성(acute) 반응과 운동훈련에 따른 장기적(chronic) 반응에 대한 결과는 일치되지 않는 다양한 결과들[12-14]이 보고되고 있어 대사적 기전은 아직까지 미흡한 실정이다. 뿐만 아니라 근력과 근비대를 위한 운동과 단백질 섭취량은 개인마다 “요구량”의 차이가 있는데, 개인의 체력수준과 훈련상태에 따라 운동강도와 운동종류 및 단백질 요구량은 개인 간 몇 배 이상의 차이를 나타내게 된다[7,8,12]. 따라서 근단백질 합성을 위한 운동과 단백질 섭취를 위해서는 다양한 조건에 대한 고려와 보다 적절한 운동방법 및 섭취량을 위한 고찰이 필요할 것으로 생각된다. 안정성 동위원소 추적체법(stable isotope tracer methodology)은 사람을 대상으로 한 대사반응에 운동 및 단백질 섭취와 관련된 근단백질 합성효과에 대한 보다 명확한 증거를 제시해 줄 수 있는 방법으로, 국외에서는 많이 활용되고 이를 근거로 한 결과가 많이 제시되고 있지만 국내에서는 많은 실험과 보고가 이루어지지 않아 아직까지 특히 운동분야에서는 접근이 어렵고 적용되지 못하고 있다.
이에 이 논문에서는 운동과 식이 섭취에 따른 골격근 단백질의 역동적인 대사를 이해하는데 큰 역할을 해온 안정성 동위원소 추적체법을 소개하고, 운동과 관련하여 골격근 단백질 대사에 영향을 줄 수 있는 요인들, 식이섭취 방법 및 종류에 따른 근단백질 합성반응의 결과들을 분류하여 논의하고자 한다. 이러한 논의는 경기력 및 건강증진을 위한 다양한 목적으로 근력과 근비대를 효과적으로 도모하고자 하는 운동선수들과 일반인들에게뿐만 아니라 건강증진을 위한 목적으로 하는 사람들에게 운동훈련과 식이섭취에 따른 골격근 단백질대사의 변화에 대한 정보를 제공할 수 있을 것이다.
본 론
1. 골격근 단백질 대사회전 측정법
인체는 지속적인 대사회전(turnover) 상태에 있으며, 골격근 단백질도 마찬가지이다. 즉, 지속적인 단백질합성과 분해과정을 겪고 있다. 근육량은 단백질합성과 분해의 상대적 속도에 의해서 결정되므로 단백질대사의 완전한 이해를 위해서는 단백질합성과 분해에 대한 이해가 함께 이루어져야 한다. 그러나 역동적인 인체의 단백질 대사회전을 특히 생명체가 살아 있는 상태(in vivo)에서 측정할 수 있는 방법은 매우 제한적이며, 이러한 측면에서 안정성 동위원소 추적체법은 가장 신뢰성 있는 측정기법이라고 할 수 있다. 이 논문에서는 단백질대사속도 분석과 관련한 대표적인 두 가지 안정성 동위원소 기법에 대해서 소개하고자 하며, 안정성 동위원소측정기법에 대해서 논의하기 전에 먼저 안정성 동위원소 추적체법을 이해하기 위해 필요한 기본적인 개념에 대해서 간략하게 설명하고자 한다.
1) 안정성 동위원소 추적체(Stable isotope tracer)
인체는 다양한 원소로 이루어져있고 각각의 원소는 하나 또는 그 이상의 안정성 동위원소를 가지는 경우가 많다. 동위원소란 양성자수는 동일하나 중성자수의 차이로 인해 발생하는 무게차이가 존재하는 동일한 원소로 화학적 성질은 차이가 없다. 예를 들어 탄소는 총 2개의 안정성 동위원소(즉, 12C과 13C)가 존재하는데, 이들은 중성자 숫자가 각각 6개 또는 7개인 차이만 존재한다. 자연계에 존재하는 안정성 탄소동위원소는 12C가 약 98.89%, 13C는 1.11% 존재한다. 시간경과에 따라서 스스로 붕괴하며 방사성에너지를 발산하는 방사성 동위원소와는 달리 안정성 동위원소는 안정한 상태로 존재하므로 인체에 유해하지 않으며, 인체내의 모든 탄소는 안정성 동위원소로 이루어져있다. 대사연구에서 가장 많이 사용되는 동위원소는 13C, 2H, 15N, 17O, 18O인데 이들은 각각의 가장 흔한(가장 가벼운) 동위원소 즉, 12C, 1H, 14N, 16O에 비해 자연계(생명체)에는 그 비율이 아주 작다[15]. 안정성 동위원소 추적체란 이와 같은 무거운 안정성동위원소(예, 13C)가 하나 또는 그 이상 표지되어 있는 생합성물질(예, 2H5-phenylalanine)을 말한다. 2H5-phenylalanine은 단백질대사 연구에서 많이 사용하는 아미노산 추적체로[16] 5개의 중수소(deuterium, 2H)가 표지된 페닐알라닌(phenylalanine)을 말한다. 안정성 동위원소를 이용한 대사연구는 이와 같은 추적체를 다양한 방법으로 체내에 주입하는데(여기서는 primed continuous infusion방법에 국한하여 설명할 것임), 주입된 추적체와 체내에 생성되어 존재하는 동일한 물질, 즉 기질(tracee)간의 질량차를 바탕으로 이들 간의 비율, 동위원소 인리치먼트(isotopic enrichment, e.g., tracer to tracee ratio)을 구하고 이를 이용해 기질의 대사속도를 추정하는 방법이다[15,17].
2) 동위원소 인리치먼트(Isotopic enrichment)
안정성 동위원소 추적체법은 일반적으로 “프라이밍 용량(priming dose)”을 주입한 후, 일정속도로 동위원소 추적체(예, 2H5-phenylalanine tracer)을 주입하는데, 체내로 주입되는 속도(infusion rate, F)와 주입된 후 체내(보다 정확하게는 특정 pool 내에서)에서 표지되지 않은 동일한 생합성물간의 비율을 통해서 특정기질의 대사속도를 추정하는 방법이다. 이때 추적체가 해당 풀(pool)에서 차지하는 상대적 비율을 동위원소 enrichment라고 하고, 가장 흔히 사용되는 동위원소 enrichment의 표현방법은 추적체에 대한 기질의 비율(tracer to tracee ratio, TTR 또는 t/T)이다[15,17]. 예를 들어, 생리학적 안정상태(a physiological steady state), 즉 공복상태에서는 혈중 기질의 농도는 일정하다. 즉, 혈중으로 들어오는 기질(tracee)의 속도(Ra, rate of appearance)와 혈류에서 제거되는 기질(tracee) 속도(Rd, rate of disappearance)가 일치한다는 것을 의미한다. 이러한 안정상태 일 때, 동위원소 추적체를 프라이밍한 후, 일정속도로 주입하게 되면 시간이 지남에 따라서 TTR이 증가하다가 어느 시점 이후부터는 고원(plateau)상태, 즉 TTR이 일정하게 유지되는 상태가 나타난다. 프라이밍의 실질적 목적은 고원에 도달하는 시간을 줄이기 위함이다. 이때를 plateau enrichment (Ep) 또는 isotopic equilibrium이라고 하고 이때는 기질뿐만 아니라 추적체가 혈중으로 들어오고 나가는 속도가 동일하게 됨을 의미한다. 다시 말해서 생리학적 안정상태 일 때, 혈중으로 들어오는 기질의 속도(Ra tracee)는 혈류에서 제거되는 기질의 속도(Rd tracee)와 동일하고 체내로 주입되는 추적체의 속도(Ra tracer, F)는 혈류에서 제거되는 추적체의 속도(Rd tracer)와 동일하게 된다. 이와 같은 관계로부터 Ra tracee (=Rd tracee)=F/Ep 공식을 유도해낼 수 있고 추정해낼 수 있다. Tracer enrichment분석은 gas chromatography-mass spectrometry (GCMS) 또는 liquid chromatography-mass spectrometry (LCMS)를 이용한다[15,17].
3) 골격근 단백질 분석방법
안정성 동위원소 추적체를 이용한 골격근 대사속도 측정법은 크게 두 가지, 즉 추적체 희석법(tracer dilution method)과 추적체 편입법(tracer incorporation method)으로 나눌 수 있으며[15,17], 단백질대사 속도는 이 두 가지 방법을 병행 이용해 추정할 수 있다. 이 두 가지 방법을 병행한 전구체-생성체관계(precursor product relation)를 이용한 근단백질 합성률(fractional synthesis rate, FSR)[18] 및 분해율(fractional breakdown rate, FBR)[19] 추정과 동정맥균형(arteriovenous balance, AV balance) 기법 및 동위원소법을 이용한 대퇴 골격근 단백질 대사속도 추정 방법은 다음과 같다.
(1)골격근 단백질 합성률(FSR)
골격근 단백질 합성속도와 단백분해속도를 측정하기 위해 가장 널리 사용되는 방법은 전구체-생성체 관계를 토대로 주입한 추적체 아미노산이 단백질로 편입되는 속도를 통해 추정하는 단백질합성률(FSR)과 Ep에 도달한 후, 추적체 주입을 멈춘 후 enrichment가 감소되는 속도(decay rate)를 통해 추정하는 단백질분해율(FBR)을 추정하는 것이다. 이 분율은 총 단백질량(total protein pool size) 중 단위시간당 새롭게 합성된 또는 분해된 단백질량(%/hour)을 의미한다.
FSR을 구하는 방법은 다음과 같다. 근단백질 FSR측정을 위해 먼저 인체 내에 아미노산 추적체(예, d5-phenylalanine)를 특정속도로 주입(primed continuous infusion)하여 Ep에 이르게 되면, 시간간격(예, 3시간)을 두고 두 번의 근육조직을 채취한다. 채취한 근육샘플을 적절하게 전 처리한 후에, 근세포질 내에 존재하는 아미노산 풀(전구체 풀)을 분리한 후, 주입한 아미노산 추적체(즉 phenylalanine)의 enrichment를 GCMS 또는 LCMS를 이용하여 분석한다. 이렇게 분석된 값을 intracellular precursor amino acid enrichment (IC precursor enrichment)라고 한다. 생리학적으로 안정상태에서의 FSR값 측정은 IC enrichment가 안정상태(steady state)임을 전제로 한다[15]. 그 다음은 근단백질을 분리하고 이를 아미노산으로 분해한 후, 추적체(phenylalanine)의 enrichment를 분석한다. 이를 protein-bound amino acid enrichment (BP enrichment)라고 한다. 이 방법을 이용해서 개별단백질(예, myosin heavy chain) [20] 또는 단백질군(예, 미토콘드리아)[21], 전체조직 단백질(mixed protein) [16,22]을 분리해서 각각의 FSR을 구할 수 있는다. FSR (%/hour) 값은 증가한 BP enrichment (변화값)을 IC enrichment 평균값으로 나누고 다시 경과한 시간(3시간)을 나눈 후 %값으로 처리하기 위해서 100을 곱하면 시간당(해당하는) 총 단백질량(pool size)의 몇 %의 단백질이 새롭게 만들어졌는지를 계산할 수 있다. 절대값(즉, 단백질합성속도)은 FSR에 총 단백질량(pool size)을 곱해서 얻을 수 있다. 지금까지는 생리학적 안정상태를 전제한 경우, 즉 공복상태 및 휴식을 취하고 있는 상태에서의 측정을 설명하였는데, 비안정상태(non-steady state)일 때, 예를 들면 영양분섭취(예, 단백질/아미노산섭취) 후 FSR 측정방법에 대해서는 좀 더 복잡한 방법이 사용된다[16,22].
(2)골격근 단백질 분해율(FBR)
근단백질 FBR 측정도 전구체-생성체 관계를 이용해 추정할 수 있다[15]. 근단백질 FBR 측정을 위해 FSR 측정에서와 마찬가지로 인체 내에 아미노산 추적체(예, d5-phenylalanine)를 특정속도(primed continuous infusion)로 주입하여 Ep에 이르게 한다. 그 후 FSR 측정법과는 달리 아미노산 추적체 주입을 중단한다. 추적체 주입을 중단하게 되면 동맥혈에서의 isotopic enrichment가 가장 먼저 감소하게 되고 IC enrichment가 감소하게 된다[15,19]. IC enrichment의 감소는 두 가지 요소에 의해서 영향을 받는데, 첫째는 혈액에서 근육세포 내로 들어오는 추적체의 상대적인 양이 줄어서이고(즉 tracee 유입이 혈액으로부터 증가하고), 두 번째는 단백질분해(추적체 주입중단에 의해서 단백질분해속도는 영향을 받지는 않는다)로 인해 출현하는 아미노산(tracee)에 의해서 IC enrichment가 줄어들기 때문이다[15]. 즉, 단백질분해속도가 크면 클수록 IC enrichment의 감소폭은 크다. 따라서 이 기법을 tracee release method라고도 한다[19]. 단백질분해는 동맥혈과 IC enrichment값, 세포질과 근단백질의 아미노산 풀 사이즈를 분석해서, 그 값을 바탕으로 FBR값을 추정할 수 있다[15]. FBR측정을 위한 실험에서 아미노산 추적체 주입을 중단하게 되면 빠른 시간 내(-1시간)에 enrichment값이 거의 바닥에 이르게 되므로, 이 enrichment가 감소하는 속도와 유형을 상대적으로 정확히 알기 위해서는 Ep에 도달한 후 한번, 추적체 주입중단 후 1시간 내에 적어도 두 번의 근육샘플, 즉 3번 이상의 근육 생검이 요구된다[15,19]. 혈액샘플은 여러 번 채취하는데 큰 어려움이 없으나 근조직을 짧은 시간 내에 3번을 한다는 것은 피검자에게나 실험자에게 부담이 된다. 이를 해결하기 위해서 개발된 기법[23]은 세 가지 서로 다른 안정성 동위원소로 표지된 동일한 아미노산추적체(예, L-[15N] phenylalanine, L-[ring-2H5] phenylalanine, L-[ring-13C6] phenylalanine)를 주입하고 Ep에 이른 후, 세 가지 다른 시점에서 주입을 중단한 후, 가장 먼저 중단한 추적체의 주입 중단시점으로부터 한 시간 후에 단 1회의 근생검을 하게 되면 하나의 근조직에서 3개의 다른 시간 때의 enrichment 값을 구할 수 있으므로 한 시간 동안의 enrichment 감소패턴을 구할 수 있다[15].
(3)동정맥균형/안정성 동위원소 추적법(Arteriovenous balance/stable isotope tracer method)
동정맥균형/동위원소추적법(A-V balance/stable isotope tracer method)을 이용한 단백질대사 연구는 운동을 포함한 다양한 조건에서 대퇴근육대사에 초점을 두고 이루어져왔으므로[24-27], 여기서는 대퇴근육의 단백질대사에 중점을 두고자 한다. 동정맥균형기법으로 대퇴근육으로 들어가는 혈류량과 동맥(모든 동맥에서는 구성성분의 농도가 일정하므로)과 대퇴정맥에서의 아미노산의 농도를 알게 되면 아미노산의 순단백질균형(net protein balance, NPB)을 구할 수 있다[15]. Phenylalanine은 근육세포 내에서 산화(oxidation)또는 합성되지 않는 필수아미노산으로 동정맥균형/안정성 동위원소추적법에서 아미노산 추적체로 널리 사용되어왔다[24-27]. 이 경우 페닐알라닌이 근세포 내로 들어가는 속도는 근단백질합성속도를 반영하고 페닐알라닌이 근세포 내에서 정맥혈로 나오는 속도는 근단백질분해속도를 반영하게 된다. 그리고 둘 간의 차가 순단백질균형(net protein balance, NPB)을 반영하게 되는 것이다. 동정맥 균형기법과 phenylalanine 동위원소 추적체를 이용할 경우, 동정맥 혈액샘플과 근조직샘플을 통해서 다양한 phenylalanine의 대사속도를 구할 수 있고, phenylalanine이 전체 근단백질에서의 구성비율(4%)[26]을 계산에 고려해 넣으면 단백질수준에서의 대사속도, 즉 NPB, 근단백질합성 및 분해속도, 근세포 내로 이동하는 속도(inward transport)와 밖으로 아미노산 이동속도(outward transport) 등을 계산해낼 수 있다(3 pool model: 동맥, 정맥, 근세포). 만약 세포간질액 샘플을 추가한다면(4 pool model)[28] 세포질과 세포간질을 구분할 수 있으므로 근세포막 수송(membrane transport rate)과 혈중에서 세포간질로의 이동속도에 대한 추가적인 정보도 얻을 수 있다. 동정맥균형/안정성 동위원소기법은 다른 많은 기법에서와 마찬가지로 장점과 제한점을 가진다. 가장 큰 장점은 FSR과 FBR이 안정상태(steady state) 일 때를 전제로 하지만[18], 이 기법에서는 안정상태가 전제되지 않으므로 음식섭취 후와 같은 비안정상태 일 때의 측정이 가능하다[15]. 또한 FSR과 FBR은 단백질합성과 분해속도를 동일한 시간 프레임에서 보기 어렵다. 그 이유는 일반적으로 FSR은 두 번의 근육 샘플링 간에 일반적으로 2시간 이상의 시간간격이 있으나, FBR 측정 시에는 1시간을 넘지 않아야 한다. 그러나 A-V balance/tracer기법은 동일한 시간프레임에서 단백질 합성과 분해를 측정할 수 있다. 반면 제한점도 있는데, 가장 큰 제한점은 동정맥에 관 삽입이 필요하므로 다른 기법에 비해 좀 더 침습성(invasiveness)의 정도가 크다는 것이다. 또한 이 기법을 사용하게 될 때, 피부 근육의 단백질대사를 볼 때에 비하면 덜 영향을 받지만, 다리 근육의 단백질대사를 볼 때에도 근조직 이외의 다른 조직(예, 지방조직, 뼈조직 등)들의 대사도 어느 정도 반영되기 때문에 순전히 다리근육의 단백질대사만을 볼 수 있다고는 할 수 없다[29]. 마지막으로 대사속도 계산에 근혈류량이 중요한 요소인데, 일정한 값을 구하기가 쉽지 않다는 것이다[29].
2. 운동훈련에 따른 골격근 단백질 대사
1) 운동종류에 따른 골격근 단백질 대사반응
저항운동(resistance exercise, RE)은 운동 후 근단백질 분해보다 근단백질 합성을 촉진하여 시간에 따른 근단백질 합성을 증가시키고, 특히 근섬유단백질인 마이오신과 액틴을 증가로 근력과 근횡단면적을 증가시키는 것으로 보고되어왔다[29,30]. 저항운동에 따른 혼합 근단백질의 합성속도(mixed muscle protein fractional synthetic rate, FSR)는 운동 후 2-3시간 내에 가장 높게 나타나며, 높은 운동량의 저항운동을 한 후에는 48시간까지도 근단백질 합성속도의 증가가 지속되는 것으로 보고된다[31]. 그러나 이러한 저항성 운동에 따른 근단백질 합성의 증가에도 불구하고 영양소 공급이 있기 까지는 단백질균형은 여전히 부적(negative)인 상태가 되는데, 이는 운동 후 손상된 근육의 재생성과 개조를 위해 더 많은 아미노산이 필요하여 단백질분해가 촉진되기 때문인 것으로 보인다[9]. Lecker et al. [32]는 공복상태에서의 저항운동 후 증가된 MPB는 근세포질(sacorplasmic pool)에서의 빠른 전환(turning over)에 의해 나타내는데, 이는 근원섬유 단백질(myofibrillar protein)의 합성을 위해 아미노산을 공급하기 위함이라고 제시하였고, 근세포질 단백질의 전환율은 근원섬유 단백질의 2배 이상인 것으로 보고되었다[33]. 즉 저항운동이 근단백질 합성을 자극하지만, 근섬유의 재형성을 위해 아미노산의 요구량이 증가하므로 추가적인 단백질의 공급없이 저항성 운동만으로는 근비대 효과를 기대하기는 어려운 것으로 보고하였다(Fig. 1).
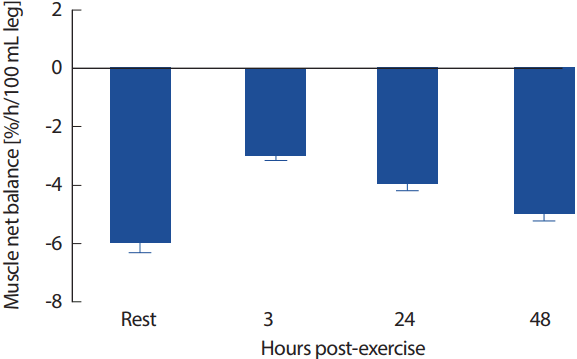
Changes in muscle net protein balance in response to resistance exercise in fasted state (adapted from [9]). In fasted state, muscle net protein balance is negative at rest. Following exercise (3 h, 24 h, 48 h), muscle net protein balance is still negative because the rate of muscle protein breakdown exceeds the rate of muscle protein synthesis. 100 mL leg: leg volume.
유산소운동(endurance exercise, EE)은 저항운동과 다르게 주로 미토콘드리아 단백질 대사회전율을 증가시키는 것으로 알려져 있다[34]. 유산소운동에 의한 미토콘드리아 밀도 증가는 산소의 이용 능력을 향상시켜 피로에 저항하는 능력을 높인다[35,36]. 저항운동훈련과 마찬가지로[37,38] 유산소운동[39,40]도 운동 후 2-4시간까지 혼합 근단백질 합성을 증가시키는 것으로 보고되고 있다.
그러나 전반적으로 유산소운동은 저항운동에 비해 전신의 근단백질 대사교환율[41]과 단백질합성[42]의 증가에 덜 효과적인 것으로 보인다. 그 이유 중 하나는 높은 에너지 사용이 요구되는 유산소운동에 따른 AMPK (AMP-activated protein kinase)의 활성이 근단백질 합성의 주요한 신호전달경로로 제시되고 있는 mechanistic target of rapamycin (mTOR) 신호경로를 억제하기 때문으로 제시되고 있다[43].
따라서 전반적으로 저항운동이 유산소운동보다는 근원섬유 단백질 합성 증가를 통해 근비대를 촉진하는 것으로 생각되지만, 저항운동은 근원섬유 단백질 합성속도를 증가시키고 유산소운동은 미토콘드리아 단백질 합성속도를 증가시켜 운동종류에 따른 골격근 단백질 대사 반응은 운동 특수성을 나타내는 것으로 보인다. 그러나 추가적인 단백질의 공급없이 이러한 운동 후 근단백질 합성을 통한 근비대 효과는 제한적일 것으로 사료된다.
2) 운동훈련 상태에 따른 골격근 단백질 대사반응
개인의 운동훈련상태는 운동 후 근단백질 합성의 증가폭과 그 증가가 지속되는 시간에 영향을 미치는 것으로 나타났다[44,45]. 장기간의 반복적인 저항운동훈련은 운동 후 근조직으로의 아미노산 공급을 더욱 원활하게 하며, 근단백질 합성과 관련된 신호전달기전을 더 민감하게 활성화시켜 근단백질 합성이 더욱 신속하게 일어나도록 하는 것으로 효과가 있는 것으로 나타났다[12,13]. 반면, 훈련된 사람의 경우는 1회성 운동 후 근단백질 합성속도의 증가가 그렇지 않은 사람들에 비해 더 빨리 안정상태로 되돌아 오는 것으로 보고되었는데, Tang et al. [14]은 장기간의 훈련으로 손상된 근육이 더 많기 때문에 손상된 근육의 재생성과 재합성을 위해서 더 많은 아미노산과 단백질이 요구되기 때문으로 제시하였다(Fig. 2).
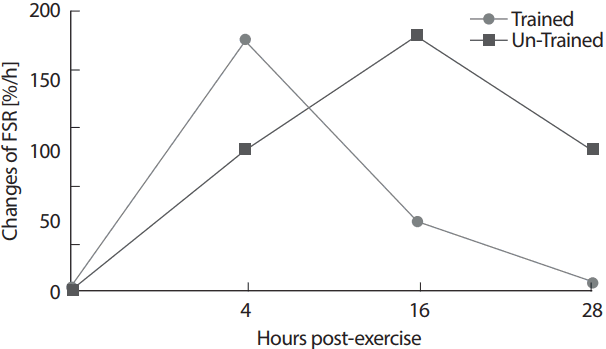
최대산소섭취량의 40% 강도에서 트레드밀 걷기운동은 훈련되지 않은 대상자의 경우엔 근단백질 합성반응을 증가시켰으나[34,42], 훈련된 수영선수들의 경우엔 고강도 수영 운동 후 근단백질 합성은 증가되지 않았다고 보고하였다[45]. 따라서 유산소운동도 훈련상태에 따라 근단백질 합성은 영향을 받는 것으로 보이며, 이러한 차이는 장기간 유산소운동 훈련에 따른 안정 시 근단백질 합성속도가 증가되었기 때문으로 제시되었다[39,40]. 반면, 최대산소섭취량의 75%에서 45분간 한쪽 다리 사이클링을 시킨 후 미토콘드리아 단백질 합성속도와 일부 근세포질 단백질(sarcoplasmic plasma)의 합성이 훈련상태와 관계 없이 훈련된 대상자과 훈련되지 않은 대상자 모두에게서 증가되었지만 근원 섬유 단백질의 증가는 나타나지 않았다고 보고되었다[46].
따라서 훈련상태에 따른 골격근 단백질 대사 반응은 장기간의 저항 운동 후에는 훈련된 대상자들의 근단백질 합성이 더 빠르게 시작되지만 지속시간은 감소되는 것으로 나타나 훈련상태에 따른 차이가 있는 것으로 나타난다. 유산소운동 후 훈련된 대상자의 근단백질 합성은 운동훈련상태에 따른 차이가 없다는 보고와 있다는 보고 등 아직 일치되지 않는 결론들이 보고된다. 그러나 이러한 변화가 운동훈련 후 안정 시 근단백질 합성속도의 증가와 관련이 있는 것으로 제시되고, 또는 일부 다른 결과들도 보고되고 있음으로 추후 더 많은 연구가 이루어져야 할 것이다.
3) 운동강도에 따른 골격근 단백질 대사 반응
운동 후 근단백질 합성의 증가는 1RM의 약 70%가 될 때까지는 운동강도에 비례(dose-response relationship)하다가 그 이상의 운동강도에서는 운동강도가 증가하여도 더 많은 근단백질 합성의 증가가 없는 것으로 제시되었다[45]. 위의 내용과 일치하게, Kraemer et al. [48]은 최대강도 70% 이상의 고강도 저항운동훈련 후 근육량의 유의한 증가를 보고했다. 이 후 많은 연구에서 근단백질 합성반응을 70-80% 1RM강도(8-12RM)에서 실시하였다[31,38,49].
그러나 최근의 연구들은 최대강도의 20%에서도 근단백질 합성반응을 자극할 수 있는 것으로 보고하는데, 저항운동 시 사지의 혈류를 제한함으로써 저강도에서도 근력증가와 근비대를 유발할 수 있다는 것이다[12,50]. 근력과 근비대가 저항운동 시 동원되는 운동신경 및 근섬유 종류와 관련이 있으며, 운동 후 근단백질 합성과 근비대는 지근섬유(Type I)에 비해 속근섬유에서 더 크게 나타나는 것으로 보고되었다[51]. 이는 혈류제한을 통해 대사적 스트레스를 더욱 증가시킴으로써 낮은 강도에서는 동원되지 않는 속근섬유(Type II)를 동원하여[52], 저강도(30-50% 1RM) 운동에서도 고강도(>70% 1RM) 운동의 근단백질 합성 효과를 냄으로써 근력과 근비대를 유도할 수 있는 것으로 제시하였다.
따라서 운동강도에 따른 골격근 단백질 대사 반응은 고강도 저항성 운동과 혈류를 제한한 저강도 저항성 운동 모두 근단백질 합성을 자극하여 근력과 근비대를 증가시키는 것으로 나타났다. 그러나 근비대는 총 운동량(수축과 부하량)에 의해 결정되며[53], 저강도 저항운동이 근비대를 유도하기 위해서는 근피로를 유발하여 속근섬유를 동원할 수 있을 만큼 충분한 자극이 주어져야 하므로[47], 이와 관련된 훈련을 계획할 때는 근단백질 합성을 자극할 수 있는 운동량에 대한 고려가 충분히 이루어져야 할 것이다.
3. 식이섭취에 따른 골격근 단백질 대사
1) 아미노산/단백질 섭취에 따른 골격근 단백질 대사 반응
안정 시 공복상태에서는 근단백질분해속도가 합성속도를 초과하여 NPB는 부적 단백질 균형상태(negative balance)에 있다. 공복상태에서 저항운동은 근단백질합성을 증가시키지만 분해도 함께 증가시키기 때문에 NPB는 여전히 부적 단백질 균형상태에 있게 된다[13]. 반면 아미노산/단백질 섭취는 근단백질 합성은 증가시키나 단백질분해의 변화는 유발하지 않으므로 NPB는 증가시키는데, 이러한 효과는 아미노산의 종류, 섭취형태, 나이, 호르몬 수준에 따라 다르게 나타난다[54]. 저항운동과 아미노산(특히 필수아미노산) 섭취를 병행할 경우, NPB를 보다 효과적으로 증가시키는데, 이는 저항운동 또는 아미노산섭취 단독 효과를 합한 수준보다 높게 나타나는 것으로 보고된다(Fig. 3).
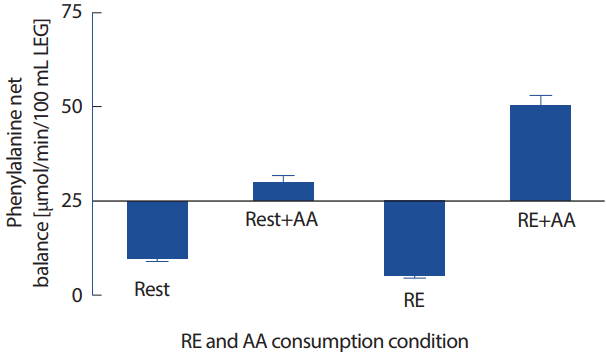
운동 후 단백질 섭취는 근손상과 이에 따른 근력약화를 억제하고 저항운동 후 손상된 근육의 회복을 촉진한다. 또한, 안정 시에 비해 저항운동은 운동 후 섭취한 아미노산을 근육이 보다 효율적으로 사용하여 근단백질의 합성을 증가시켜 근육량과 근육의 크기를 증가시킬 수 있는 것으로 제시되었다[53,54]. 이와 같은 아미노산의 섭취에 따른 NPB는 아미노산섭취량에 비례하여 증가하는 것으로 보인다[57,58].
또한, 탄수화물에 추가적인 단백질과 필수아미노산의 섭취는 저항운동 후 탄수화물만을 섭취한 경우보다 근단백질 합성율을 증가시키는 것으로 보고되었다[59-62]. 단백질/필수아미노산섭취에 추가적으로 탄수화물섭취 할 경우 인슐린을 증가시키고 이는 저항운동 후 근단백질 분해를 감소시키는 역할을 한다. 근단백질 분해가 감소된 상태에서의 혈중 높은 수준의 아미노산은 근단백질 합성을 증가시키므로 결과적으로 양적(positive) 단백질 균형상태를 나타내게 된다[60-62].
따라서 공복상태에서의 저항운동은 근단백질합성과 동시에 근단백질 분해를 증가시켜 근단백질 균형을 감소시키지만, 아미노산의 추가적인 섭취는 근단백질 균형을 증가시키고 근력과 근비대를 촉진하는 효과가 있는 것으로 나타났다. 이러한 효과는 섭취한 아미노산의 양에 비례하여 나타나게 되므로 저항운동과 아미노산의 섭취를 병행하는 것이 동화반응을 촉진하는데 효과적일 것이다.
2) 단백질 섭취량과 종류에 따른 골격근 단백질 대사 반응
여러 선행연구들에서 필수아미노산(essential amino acid, EAA)만이 근단백질 합성 촉진의 생리적 작용을 하는 아미노산으로 제시하였다[61-63]. 필수아미노산의 섭취는 혈중 아미노산의 농도를 빠르게 증가시키고, 이것은 비필수아미노산(non-essential amino acid)보다 근단백질 합성을 자극한다[61]. 또한 안정 시와 운동 후 모두 필수아미노산의 양에 비례하여 근단백질 합성을 촉진하는 것으로 보고되었다[33, 63].
최근 Moore et al. [66]은 저항성 운동 후 근단백질 합성을 최대로 증가시키기 위해 필요한 필수아미노산 섭취량에 대한 연구를 수행했다. 이 연구에서 저항성 운동 후 피검자들은 0, 5, 10, 20, 40 g의 단백질을 섭취하였는데, 근단백질 합성속도는 20 g의 단백질 섭취량(필수아미노산 8.6 g 포함)까지 섭취량에 비례하여 증가하였으나 그 이후로는 더 이상의 합성효과는 없었다. 따라서 저항성 운동 후 근단백질 합성을 최대화하기 위해서는 8-10 g의 필수아미노산 섭취가 필요할 것으로 제시하였다.
근단백질 합성반응은 섭취 후 단백질이 소화되는 속도에 의해서도 영향을 받는 것으로 보인다[67]. 빠르게 소화되는 단백질은 혈중 아미노산농도를 빠르게 증가시키고 전신 단백질 합성을 증가시키는 효과가 있는 반면, 느리게 소화되는 단백질은 혈중 아미노산농도를 천천히 증가시키고 이는 주로 전신 단백질분해를 억제하는 효과를 나타내는 것으로 보고 되었다[68]. 이러한 단백질 소화속도에 따른 차이에 대해 Burd et al. [69]은 안정 시와 저항성 운동 후 웨이 단백질이 카제인 단백질이나 콩 단백질보다 근단백질 합성에 더욱 효과적인 것으로 보고하였다. 이는 웨이 단백질이 다른 단백질보다 근육단백질의 주성분인 필수아미노산 류신을 더 많이 포함하고 있으며, 빠르게 소화되기 때문에 근단백질 합성을 촉진시키는 것으로 제시하였다. Hartman et al. [70]은 우유 단백질이 저항운동 후 콩 단백질보다 근단백질 합성을 촉진시키는 것으로 보고하였다. 그러나 Tipton et al. [7]은 웨이와 카제인 단백질 모두 저항운동 후 순 근단백질 균형을 동일하게 증가시킨다고 보고하였다. 이는 카제인 단백질이 빠르게 흡수되는 웨이 단백질보다 저항운동 후 근단백질 분해를 더 많이 억제하기 때문인 것으로 제시하였다.
이와 같이 단백질 소화속도에 따른 근단백질 합성반응은 일치되지 않는 결과들이 보고되는데, Paddon-Jones et al. [72]은 장에서 천천히 소화되는 단백질은 내장에 흡수가 많이 되어 말초로 순환하는 혈액의 아미노산 농도를 증가시키지 못하기 때문에[73] 빠르게 소화되는 단백질에 비하여 근단백질 합성반응이 낮다고 제시하였다.
요약하자면, 저항성 운동 후 근단백질 합성은 단백질섭취량이 20 g(필수아미노산 8-10 g을 포함)이 될 때가지 단백질섭취량에 비례하여 증가하는 것으로 보이며, 빠르게 소화되는 단백질이 느리게 소화되는 단백질에 비해 근단백질 합성이 보다 효과적으로 촉진되는 것으로 보인다. 그러나 아직까지 일치되지 않는 결과들이 보고되므로 이와 관련된 추가적인 연구들이 필요할 것이다.
3) 단백질의 섭취 시기에 따른 골격근 단백질 대사 반응
운동 전과 후의 단백질 섭취시기는 오랫동안 저항운동훈련을 하는 선수들과 연구자들에게 근비대를 위한 중요한 요인으로 고려되어 왔다. 그러나 운동전과 후의 아미노산 섭취에 따른 근단백질 합성효과는 일치되지 않는 결과들이 보고되고 있다. Tipton et al. [74]은 단백질섭취 시기가 운동 전과 후에 관계없이 근단백질합성을 유사하게 증가시켰다고 보고 하였다. Rasmussen et al. [75]도 운동 후 1시간과 3시간 후 근단백질 합성속도를 비교하였으나 차이가 없다고 하였다. Hartman et al. [70]은 운동 직후와 2시간 후에 단백질섭취를 한 경우를 비교한 결과, 운동 직후 섭취했을 경우 근육량과 근섬유 비대가 더 크게 증가한 것으로 보고하였다.
반면, Wolfe [76]는 운동 전 6 g 필수아미노산과 35 g의 포도당을 혼합해서 섭취한 경우 운동 직후 섭취하는 것보다 운동하는 동안 근육으로 아미노산의 전달과 흡수를 증가시켰으며, 운동 후 1시간 동안 더 큰 근단백질 합성 반응을 촉진하여 순 근육단백질 균형이 더 많이 증가했다고 보고하였다[61]. 이에 상응하게 Esmarck et al. [77]은 노인을 대상으로 한 연구에서, 운동 직후 단백질/탄수화물/지방이 포함된 혼합음료를 섭취한 경우에 비해 2시간이 지난 후 섭취한 경우 근력이 덜 증가하였다고 보고하였다.
이와 같이 단백질 섭취시기에 대한 근단백질 합성효과는 아직까지 연구결과들이 일치되지는 않고 있지만, 운동 후 가능한 빠른 시간 내에 단백질을 섭취하는 것이 근육으로 아미노산의 전달과 흡수 및 이용을 통한 근단백질 합성을 증가시키는 것으로 제시되고 있다[69].
4) 훈련상태에 따른 단백질 섭취량
북미에서는 일반인들의 단백질 섭취권고량(recommended dietary allowance, RDA)은 0.8 g/kg/day이지만[78], 저항성 운동 훈련을 수행하는 운동선수나 규칙적인 운동을 하는 사람들의 단백질 섭취량은 좌업 생활을 하는 사람들보다는 더 많아야 할 것으로 제시하고 있다. 이는 장기간의 저항운동 훈련동안 제지방량과 근비대를 위해서는 순 단백질 균형이 증가된 상태가 유지되어야 하는데[71], 저항운동, 특히 신장성 운동 후, 손상된 근육을 회복하기 위해서는 더 많은 단백질이 요구되므로 선수들은 제지방량을 증가시키기 위해서 단백질 섭취를 증가시켜야 한다는 것이다[79,80]. 또한, 고강도로 훈련된 파워리프터와 보디빌더와 같이 높은 근육량을 가진 사람들은 안정 시 단백질 대사전 환율이 높기 때문에 좌업생활인보다 단백질 요구량이 증가하고, 순 근단백질 균형을 양적(positive)으로 유지하기 위해서는 1일 1.6-1.8 g/kg을 섭취해야 한다고 권고하고 있다[81].
Tarnopolsky et al. [82]은 저항성 운동훈련을 하는 사람들에게 추가적인 단백질 섭취는 훈련초기에 근비대를 유도한다고 하였고, Tipton et al. [9]은 저항운동 후 단백질 섭취는 근단백질 분해를 감소시키고 근단백질 합성을 증가시켜 순 단백질 균형을 보다 효과적으로 증가시킨다고 보고하였다. 그러나 저항성 운동 시 훈련된 선수들에게 단백질 섭취가 추가적인 효과를 가져온다는 보고[62,83]와 효과가 없다는 보고[84,85] 등 일치되지 않는 결과들이 보고되는데, Kraemer et al. [58]은 훈련된 사람들에게 며칠간의 단백질 보충제 섭취는 훈련 양이나 강도와 상관없이 저항운동의 효과에 영향을 주지 않지만, 장기간 동안의 보충제 섭취는 효과를 증가시킨다고 하였다.
이와 같이 규칙적으로 운동하거나 훈련된 선수들은 그렇지 않은 사람들에 비해 안정 시 단백질 대사전환율(합성과 분해)이 높고, 근 손상을 회복하기 위해 더 많은 단백질 섭취가 필요한 것으로 보인다. 그러나, 단백질요구량은 운동강도, 기간, 빈도, 그리고 개인의 훈련상태에 따라 다르므로 이를 고려하여 단백질을 섭취함으로써 효과적으로 저항훈련효과를 증대시킬 수 있을 것으로 사료된다.
결 론
이 연구는 운동과 관련하여 골격근 단백질 대사에 영향을 줄 수 있는 요인들, 식이섭취 방법 및 종류에 따른 근단백질 합성반응을 안정성 동위원소 추적체법을 이용한 연구결과를 고찰한 결과 다음과 같은 결론을 내렸다. 저항운동은 근원섬유 단백질 합성속도를 증가시키고 유산소운동은 미토콘드리아 단백질 합성속도를 증가시켜 운동종류에 따른 골격근 단백질 대사 반응은 운동 특수성을 나타내며, 운동훈련상태와 운동강도에 따른 차이가 있는 것으로 보인다. 그러나 공복상태에서 저항운동은 근단백질합성을 증가시키지만 분해도 함께 증가시키기 때문에 여전히 부적 단백질균형상태에 있게 되므로 저항운동과 아미노산의 섭취를 병행할 경우 보다 효과적인 것으로 보인다. 단백질요구량은 운동강도, 기간, 빈도, 그리고 개인의 훈련상태에 따라 차이를 나타내며, 단백질의 질(즉, 상대적인 필수아미노산의 양)과 나이 등에 의해서도 영향을 받으므로, 이러한 요소들을 고려하여 단백질을 섭취함으로써 효과적으로 저항훈련효과를 증대시킬 수 있을 것으로 사료된다. 또한 성별에 따라서도 근단백질 합성반응은 달라지는데 아직까지 다양한 결과들이 보고되고 있으며 훈련량에 따른 결과도 일치되지 않는 결과가 보고되고 있고, 노화와 질병에 따른 연구에 비하여 운동선수들에 대한 연구는 부족한 실정이므로 이와 관련된 추가적인 운동과 단백질 섭취에 관한 연구가 이루어져야 할 것으로 제시하는 바이다.