청소년 사이클리스트의 최대 스프린트 사이클링 중 에너지 수요 기여도 분석
Energy Demands during Maximal Sprint Cycling in Youth Cyclists
Article information
Trans Abstract
PURPOSE
The purpose of this study was to determine all different energy systems contribution during the maximal sprint cycling in youth cyclists.
METHODS
Fifteen male and female youth cyclists participated in this study. A simulated maximal 10 sec maximal sprint cycling was carried out. During maximal sprint cycling, energy contribution (phosphagen, WPCR; glycolytic, WLa -; oxidative system WOXI in kJ and %), maximal lactate production rate (VLamax), difference of lactate (ΔLa-), highest lactate (Peak La-), peak power (Wpeak) and anaerobicalactic time (tAlac) were analyzed using off VO2 kinetics, trapezoidal method, O2-lactate equivalent, and caloric quotient.
RESULTS
During the maximal sprint cycling, WPCR was significantly higher than WLa - and WOXI (p=.008; d: 0.98, p<.001; d: 2.79, respectively). Also, WLa - was significantly higher than WOXI (p<.001; d: 2.56). Additionally, low negative and moderate positive correlations were observed between tAlac, Wpeak and WPCR in kJ (r=.43, r=.63, respectively). Furthermore, high and moderate positive correlations were calculated for ΔLa-, Peak La-, VLamax and WLa - (r=.91, r=.86, r=.68, respectively).
CONCLUSIONS
Phosphagen and glycolytic energy systems were predominantly used during maximal sprint cycling. Therefore, physical training program for cycling athletes should be directed to the development of anaerobic metabolism because anaerobic performance is crucial during the short term cycling sprint phase.
서 론
육상 남자 800 m 트랙경기, 1,500 m 스피드 스케이트 또는 경륜경기의 경우 약 120초 이내에 마쳐야 하는 경기이다. 잘 훈련된 800 m 스프린터의 경우 경기 중의 에너지 시스템 기여도는 유산소성 에너지가 약 60-70%, 무산소성 에너지가 30-40%인 것으로 보고되고 있다[1,2]. 따라서 800 m 트랙경기와 유사한 경쟁 스포츠 종목의 경우 운동 수행 시의 각기 다른 두 에너지 시스템의 중요 기여도를 알 수 있다. 경기의 우승자를 가리는 중요한 운동능력 요인 중 하나로 결승지점에서의 마지막 전력 질주 즉, 최종 스퍼트를 들 수 있다. 선수들은 이 구간에서 최대의 운동능력을 발휘하기 위해 이전까지는 호기성 에너지를 더 많이 활용하여 혐기성 에너지의 근원을 확보하는 것이 경기의 전략적 요인 중 하나이다[3].
이와 관련하여 근수축 시 아데노신 삼인산(adenosinetriphosphate: ATP)의 수요가 미토콘드리아 호흡을 초과하면 대사 산증을 유발하는 수소이온(H+)의 축적이 발생한다[4]. 대사 산증은 세포의 pH 농도 저하로 유발되며, 해당과정의 중요 효소인 포스포프룩토키나아제(phosphofructokinase, PFK)를 점차적으로 억제한다. 따라서 이 수준의 운동강도는 제한적일 수 있다. 젖산 축적은 이러한 세포성 산증과 시점 일치성 때문에 세포 혐기성 대사분석의 좋은 간접적 변인으로 간주되고 있다. 그러므로, 젖산농도는 서로 다른 에너지 공급 시스템의 사용을 나타낼 수 있는 척도로 쓰인다[4]. 생리학적 반응으로 혈중 젖산농도, 최대 산소섭취량(VO2max), 단일지수 모델의 빠른 구성요소(VO2 fast component) 그리고 off VO2 kinetics는 운동부하 특성과 관련이 있으며, 또한 활성화된 근육량, 자세와 모터 패턴 등에 의해 다르게 반응한다[5]. 또한, 생리학적 효과 및 반응은 최대 운동부하 동안 더욱 높게 나타나는 것으로 보고되고 있다[6]. 따라서, 유무산소 운동능력 분석을 위해서는 VO2max 그리고 최대 누적 산소 결핍(maximal accumulated oxygen deficit, MAOD) 등이 널리 사용되는 대사 분석 지표로 활용되고 있다[7]. All-out 스프린트 테스트는 선수의 최대 스프린트 운동능력(예: 최대 출력)을 평가하고 젖산을 교환 및 제거하는 능력을 설명해주며 해당 선수의 무산소성 운동능력을 가늠할 수 있는 척도가 되고 있다[8,9]. 최대 해당과정 파워(glycolytic power)는 초당 최대 젖산 생성률(maximal lactate production rate, VLamax)로 설명할 수 있으며, 운동선수의 생리학적 무산소성 운동능력 프로파일에 대한 더 넓은 해석을 가능하게 한다[9-12].
이러한 선행연구들에서는 무산소성 운동능력을 검사하기 위해 15 seconds 최대 스프린트 사이클링을 실시하였다[10,12,13]. Heck et al. [9]의 연구에서는 VLamax 분석 시 최대 운동부하 시간을 10초 이내로 제한한다. 최대 운동부가 길어질수록(10초 이상) 대사(혹은 호흡) 산증으로 인하여 PFK가 억제되어 젖산 생성률 속도가 감소하기 때문이다. 또한, VLamax 변인만으로는 다른 에너지 대사 시스템인 포스파젠 과정 그리고 유산소성 에너지 시스템 소비 기여도를 스프린트성 최대 운동부하 도중의 에너지 대사의 과정을 모두 설명할 수 없다는 단점이 있다. 이에 반해 Beneke et al. [14]의 모든 에너지 대사 비율 계산법은 운동 또는 실제 경기 도중 시행할 수 있는 측정방법이다. 또한, 실제 경기력 분석에 있어 가장 근접한 시뮬레이션을 통하여 모든 에너지 대사 수요의 기여도를 분석할 수 있어 최근 태권도, 유도, 복싱, 가라테, 농구, 탁구, 2,000 m 조정경기, 실내 암벽 등반 등과 같은 경기종목에서 선수의 운동수행력(경기력)을 분석하는 데 이용되고 있다[14-21]. 하지만 경륜 등과 같이 혐기성, 호기성 에너지 시스템의 혼용사용이 나타나는 자전거경기의 경우 이러한 연구를 통한 에너지 기여도에 대한 탐색은 충분하지 않다.
따라서 본 연구의 목적인 선행연구들에서[10,12,13] 사용되고 있는 분석방법을 최대 10초 스프린트 사이클링 운동에 적용하여 포스파젠 시스템, 해당과정 시스템, 그리고 산화적 인산화 시스템과 같은 세가지 에너지 대사의(Fig. 1) 기여도를 비교 및 분석하고자 하였다.
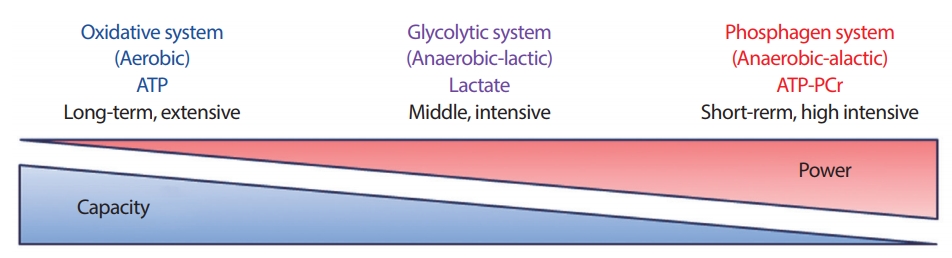
Illustration of three energy system regard to exercise duration and intensity.
연구 방법
1. 연구 대상
본 연구의 대상자는 독일 Nordrhine-Westphalia 주에 속해있는 엘리트 사이클링 남녀 청소년 엘리트 선수 15명(남: 10, 여: 5)을 대상으로 실시되었다. 실험 전 피험자들은 연구자로부터 위험요소들과 연구의 목적 및 내용에 대한 설명을 충분히 숙지한 후 실험에 자발적으로 참여하였다. 본 연구는 독일체육대학교 쾰른 연구윤리위원회 심의(No. 51/2017)를 통과하였으며, 적용된 프로토콜은 헬싱키 선언과 일치하였다(with the Declaration of Helsinki). 본 연구대상자의 신체적 특성은 Table 1과 같다.
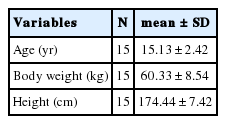
Anthropometric data
2. 연구절차
1) 신체조성
신장은 신장계(SECA 213, GmbH & Co. KG, Germany)를 이용하여 직립한 표준자세로 측정되었다. 체중은 체중계(BG-17 Beurer, Söflinger, Germany)로 측정되었다.
2) 최대 운동부하 10초 사이클링 테스트
모든 스프린트 테스트는 SRM Ergometer (Schoberer Radmesstechnik, GmbH, Germany)를 이용하여 실행되었다. 좌석 및 핸들바의 높이는 모든 테스트에서 각 피험자마다 맞춰 동일하게 유지 되었다. 본 스프린트 테스트 전 50 W에서 10분간 준비운동을 실시하였다. 준비운동 직후, 5분간 수동적 휴식을 취하였다. 본 실험 전 안정 시 혈중 젖산 농도를 측정하기 위해 귓볼 모세혈관에서 20 μL 혈액샘플을 채취하였다. 3초의 카운트다운 후, 피험자는 착석 위치에서 10 seconds all-out 스프린트 테스트를 케이던스 130 rpm으로 최대한 페달을 밟기 시작하였다[9,12]. 10초 테스트 후, 10분간 분당 최대 혈중 젖산농도를 분석하기 위해 채혈을 실행하였다(Fig. 2). 혈중 젖산 농도분석을 위하여 효소 전류측정 센서 칩 시스템 기반의 젖산 분석기(Biosen S-line, EKF diagnostic salse, GmbH, Germany)를 사용하였다. 산소 섭취량(VO2)은 Breath-bybreath (Oxycon Mobile, CareFusion, Germany) 측정방법을 통하여 데이터값이 산출되었으며(안정 시 5분, 10초 스프린트 사이클링, 운동 후 산소 소비: off VO2 6분) 호흡 가스 분석기의 gas calibration (16% O2, 5% CO2; RiessnerGase, Germany) 그리고 Turbine volume transducer는 3 L syringe (Hans Rudolph Inc., USA)를 사용하여 실행되었다.
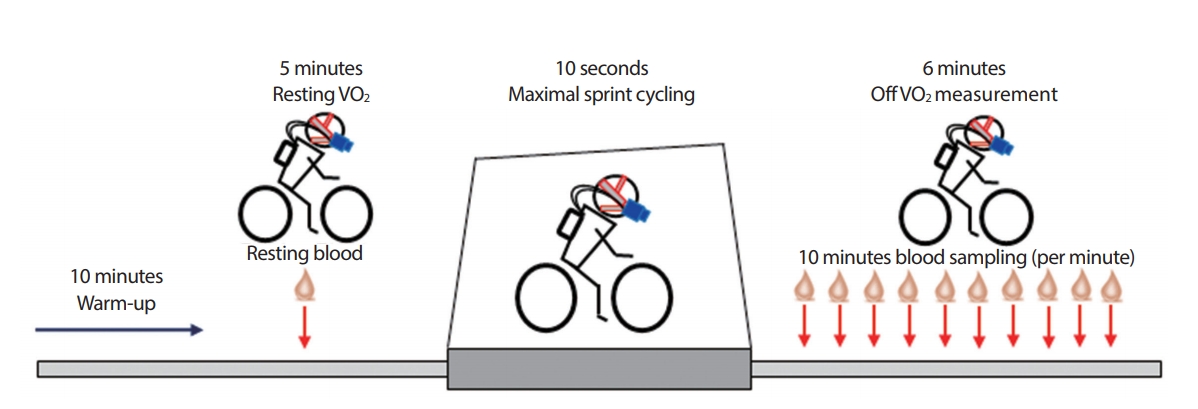
Illustration of study design.
3) 초당 최대 젖산 생성률(VLamax)
VLamax는 최대 해당과정 운동능력 검증을 위해 추가로 산출되었다[9]. 스프린트 운동 부하 후 최대치 그리고 안정 시 혈중 젖산의 차이를 통해 산출되었다. 데이터 산출에 있어 최대 스프린트 운동 부하의 초반부에 지배적으로 쓰이는 포스파젠 에너지 시스템 소비 시간을 산출하기 위해 Peak watt 데이터의 3.5% 저하되는 지점을 포스파젠 무산소성 운동시간(tAlac)으로 지정하였다[10,12,13]. VLamax의 공식은 다음과 같다 Eq (1):
VLamax=(Lamax–Lar) / (tExer–tAlac)
VLamax=초당 최대 젖산 생성률
Lamax=혈중 최대 젖산 값
Lar=안정 시 혈중 젖산 값
tExer=총 운동부하 시간
tAlac =포스파젠 무산소성 운동시간
4) 에너지 시스템 수요 기여도 분석
포스파젠(WPCR), 해당과정(WLa-) 그리고 산화적 인산화(WOXI) 에너지 시스템 분석은 운동 중 산소 섭취량(VO2), 최대 혈중 젖산 농도 및 운동 후 초과산소 섭취량(EPOCFAST)에 의해 산출되었다[14,15]. WOXI 에너지는 VO2에서 VO2REST를 뺀 곡선의 면적을 조각으로 나눈 후 사다리꼴 적분 근사식 방법(trapezoidal method)을 통하여 산출되었다[15-17,22]. VO2REST 는 테스트 전 5분간 자전거 에르고미터에서 앉은 자세로 측정되었으며 그중 마지막 30초를 기준으로 결정되었다. WPCR 시스템은 10초 스프린트 테스트 후 초과산소 섭취량(EPOCFAST)을 고려하여 산출되었으며, off VO2의 단일 및 이중 지수모델(mono- and bi-exponential models)에 맞춰 분석되었다. 이중 지수모델의 느린 구성 요소(slow component)는 무시되었으며 단일 지수모델의 빠른 구성 요소(fast component)가 WPCR 의 분석에 쓰였다[14-17,23]. WLa- 시스템 분석은 1 mmol·L-1 혈중 젖산 축적의 동등 값인 체질양의 3 ml O2 kg-1을 사용하였다[24]. 열량 지수 20.92 kJ (caloric quotient)는 모든 에너지대사 시스템 분석에 사용되었다[25]. 혈중 젖산 차이 값(ΔLa-)은 운동 부하 후 최대 젖산 축적 값에서 운동 부하 전 안정 시 데이터를 뺀 값으로 분석되었다. 총 에너지대사 수요(WTOTAL)는 세 가지 에너지대사 시스템의 합으로 산출되었다(WPCR+ WLa-+WOXI in kJ)[14].
3. 자료처리방법
본 연구에서 측정된 자료는 윈도우용 GraphPad Prism ver 8로 분석하였다. 측정 변인들의 정규성 분포를 확인하기 위해 샤피로-윌크 검정(Shapiro-Wilk)을 실시하였다. 각 에너지 시스템 간의 유의성 검증은 일원배치 반복측정 분산분석(one-way repeated-measure analysis of variance)으로 하였으며, 변인 간 유의한 차이가 있을 경우 Bonfferoni를 통해 사후 검정을 실시하였다.
본 연구의 모든 유의수준(α)은 p<.05로 설정하였다. 효과 크기를 분석하기 위해 Effect size Cohen’s d를 실시했으며, 효과 크기는 small (0.2), moderate (0.5), 그리고 large (0.8)로 각각 설정되었다[26]. 추가적으로 변인 간의 상관분석을 위해 Pearson correlation을 실시하였다.
연구 결과
1. 에너지 시스템 그리고 생리학적 변인
Table 2에 나타난 바와 같이 WPCR과 WLa- 에너지 시스템에서 통계적 유의한 차이를 나타냈다(p = 0.008; d: 0.98). WPCR 그리고 WOXI 에너지 시스템에서 유의한 차이를 나타냈다(p<0.001; d: 2.79). 또한, WLa- 그리고 WOXI 시스템에서도 통계적 유의한 차이를 나타냈다(p <0.001; d: 2.56) (Table 2).
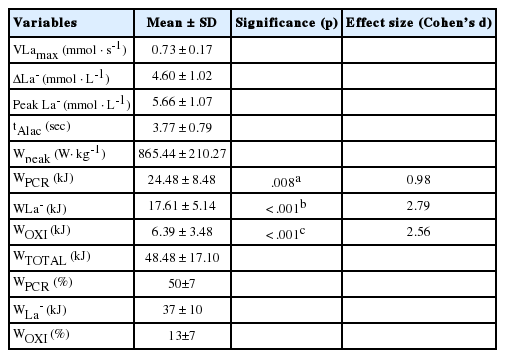
Energy contribution and other physiological parameters
WPCR in kJ와 tAlac, WPEAK 변인 간 각 부적상관 그리고 정적상관성(r= 0.43, CI: -0.76-0.08, p = 0.094; r= 0.62, CI: 0.17-0.85, p = 0.011)이 있었으며, WLa- in kJ 그리고 ΔLa-, Peak La-, VLamax 변인 간 각 유의한 정적상관성(r= 0.91, CI: 0.74–0.97, p <0.001; r= 0.86, CI: 0.64–0.95, p <0.001; r= 0.68, CI: 0.28–0.88, p = 0.004)이 있는 것으로 나타났다(Figs. 3, 4).
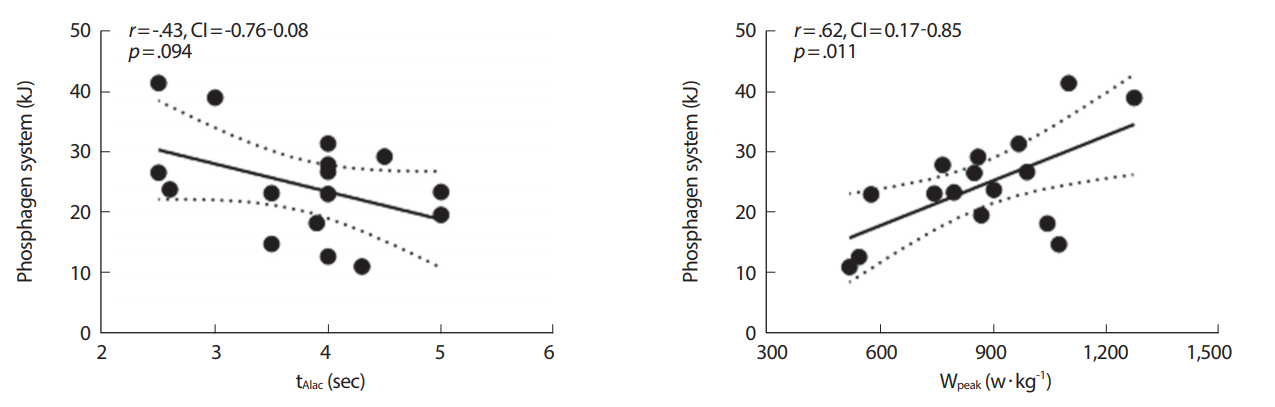
Relationships between anaerobic-alactic time (tAlac), highest watt (Wpeak) and phosphagen system in kJ (WPCR). The linear fits are significant (p<.05) in kJ (WPCR). The linear fit is significant (p<.05).
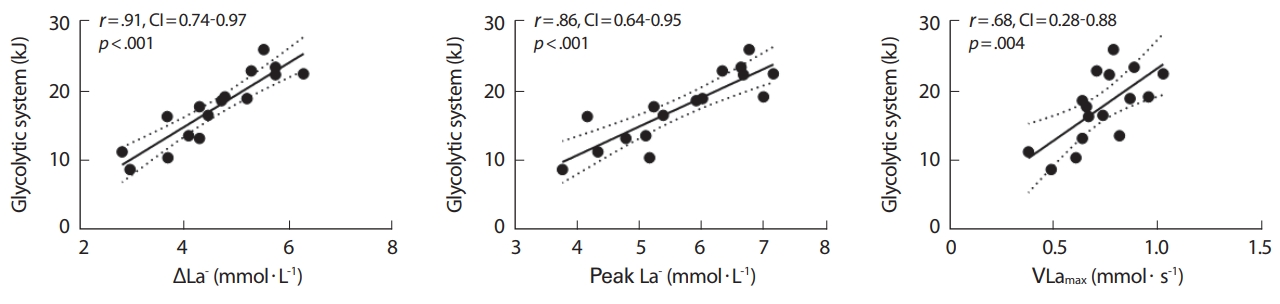
Relationships between difference of lactate (ΔLa-), highest lactate (Peak La-), maximal lactate production rate (VLamax) and glycolytic system in kJ (WLa-). The linear fits are significant (p<.05).
논 의
본 연구는 기존의 VLamax 분석을 통해서는 명확하게 구분하기 어려웠던 경기 중 에너지 대사 기여율을 모든 에너지 대사 시스템의 비율법을 이용하여 조사하였다.
본 연구의 주요 결과로서 최대 10초간 스프린트 사이클링 운동부하 중 포스파젠 에너지 시스템 수요도가 지배적으로 쓰이는 것으로 나타났으며 해당과정 및 산화적 인산화 시스템은 비교적 낮은 비율을 차지하는 것으로 분석되었다. 각 에너지 시스템은 통계적으로 유의한 차이를 나타냈으며 생리학적 변인(tAlac, WPEAK, ΔLa-, Peak La-, VLamax) 및 포스파젠 그리고 해당과정 에너지 시스템 간 각 유의한 부적 및 정적 상관성을 보였다.
기존 선행연구자들의 주장 또한 100 m 단거리 최대 운동부하 시 지배적인 에너지 대사 수요 기여도는 크레아틴 인산(creatine phosphate: PCr)의 근육 저장고에 많은 부분을 의존하는 포스파젠 시스템으로 알려지고 있다[27,28]. 포스파젠 시스템에서 크레아틴 키나제(creatine kinase, CK)는 PCr을 크레아틴으로 분해하고 PCr에서 아데노신 이인산(adenosine diphosphate, ADP)으로 무기 인산(Pi)을 전달하여 ATP를 형성한다. 따라서 포스파젠 시스템이 작동하는 동안 ATP는 매우 높은 속도로 재생될 수 있으며 크레아틴 인산이 남아있는 한 근육 ATP는 적당히 일정한 수준을 유지될 수 있다고 보고되었다[27,28]. 본 연구에서는 포스파젠 에너지 시스템이 사용된 시간은 최대 10초 운동부하동안 초반부 평균 3.77초인 것으로 분석되었으며, 포스파젠 에너지 분석과 부적 상관성을 나타냈다. 이러한 결과는 최대 스프린트 운동부하의 다른 선행연구에서도 유사한 결과로 뒷받침된다[12,13]. 하지만 본 연구의 최대 10초 운동부하 시 무산소성 구성요소, 특히 포스파젠 시스템 고 에너지 인산의 사용 비율은 계산된 것보다 적을 수 있다. 왜냐하면 최대 운동부하 후 VO2의 빠른 구성요소(fast component)는 WPCR의 상환뿐만 아니라 산소(oxygen) 저장고의 보충을 나타낼 수 있기 때문이다[29]. 해당과정 에너지 대사 시스템의 근본적인 연료 공급원으로 포도당(glucose)에 의해 작용하며 스프린트와 같은 폭발적인 최대운동부하 중 근육 ATP 회전율에 기여하게 된다. 이전의 선행연구에서는 10초간 지속하는 스프린트 운동 중에 해당과정 에너지 시스템 기여도는 최대 55% 이상을 차지할 수 있다고 한다. 그러나 본 연구결과에서는 혈중 젖산 축적 농도로 분석된 해당과정 시스템의 기여도는 37%, 산화적 인산화 시스템은 13%로 나타났다. 이러한 연구 결과 차이는 기존 선행연구 대부분은 누적 산소 결핍 및 포스파젠/해당과정 에너지를 합친(PCr/La total) 계산법으로 호기성 에너지와 혐기성 에너지를 분석하는 것처럼 에너지 시스템의 기여도를 분석함에 있어 다른 방법을 채택하는 것에 의한 것으로 생각된다[30,31]. 다른 한편으로는 포스파젠 및 해당과정 에너지 시스템 수요 기여도는 피험자의 근육량에 따라 큰 차이를 나타낼 수 있다[9,11,28]. 남녀 청소년 사이클링 선수들은 상대적으로 성인 선수들에 비해 근육량이 낮으므로 앞서 언급한 차이를 유발할 수 있는 것으로 보여지며 이와 관련한 추후 연구가 이루어져야 할 것이다.
Quittmann et al. [13], Hauser et al. [12]의 연구에서는 최대 스프린트 사이클링 후 VLamax 값이 평균 0.63-0.91 mmol·s-1으로 보고되었으며, 본 연구의 VLamax 역시 평균 0.73 mmol·s-1으로 유사한 결과를 보였다. 그뿐만 아니라 VLamax와 해당과정 에너지 시스템은 정적 상관성이 나타났다.
결 론
본 연구 결과를 종합해 보면 청소년 엘리트 사이클 선수들의 최대 10초 스프린트 사이클링 운동부하 시 모든 에너지 대사 시스템 중 포스파젠 시스템이 운동 초반부에 지배적으로 사용되었으며, 해당과정 에너지의 기여도 또한 중요한 것으로 나타났다. 반면에, 산화적 인산화 에너지 대사는 비율적으로 큰 부분을 차지하지 않았다. 이러한 사실을 토대로 마지막 결승지점에서의 폭발적인 운동능력을 필요로 하는 사이클 선수들의 훈련 프로그램 구성에 있어서 포스파젠 및 해당과정 에너지 시스템을 향상시킬 수 있는 훈련에 중점을 둔다면 경기력 향상에 도움이 될 수 있을 것이다.
Notes
이 논문 작성에 있어서 어떠한 조직으로부터 재정을 포함한 일체의 지원을 받지 않았으며, 논문에 영향을 미칠 수 있는 어떠한 관계도 없음을 밝힌다.