골격근 위축에 대한 운동 메커니즘: 기전적 분석
Role of Exercise in Skeletal Muscle Atrophy: A Mechanistic Investigation
Article information

Trans Abstract
PURPOSE
Skeletal muscle atrophy induces overall health problems and many related diseases in older adults. In particular, sarcopenia is related to lowered quality of life, decreased physical activity, high levels of morbidity and mortality owing to chronic diseases, and even falling. Despite the many clinical studies on skeletal muscle atrophy, a little study had been about the exact mechanism of skeletal muscle atrophy at the molecular level. In 2017, A disease code (ICD-10-CM) for skeletal muscle atrophy related to sarcopenia was announced to attempt a clinical approach in the United States. According to these approaches, non-invasive clinical treatment is the most effective method for treating skeletal muscle atrophy. The purpose of this study was to analyze the molecular mechanisms of muscular exercise and related skeletal muscle atrophy factors.
METHODS
This systemic review focused on skeletal muscle atrophy and muscular exercise. The keywords were used on “MeSH: muscle atrophy OR skeletal muscle atrophy OR muscle OR atrophy AND physical exercise OR exercise OR exercise training” for English and Korean. This paper searched PubMED, OVIDMEDLINE, and EMBASE for literature consideration.
RESULTS
Skeletal muscle atrophy was related to a complex molecular network, and exercise affects IGF-1/Akt/mTOR signaling. The related skeletal muscle atrophy factors were evaluated as MurF1, MAFbx, IGF-1, and NFkB. We proposed new related factors such as ATF 4, Gadd45a, and p21; however, the results related to exercise were not shown in recent studies.
CONCLUSIONS
In conclusion, we identified skeletal muscle atrophy factors at the molecular level of muscle physiology, and these new factors may become an interesting field of study in clinical human trial and animal studies.
서 론
영양부족, 근육감소, 노화 그리고 운동부족으로 인한 골격근 위축은(muscle atrophy) 건강과 관련되는 여러 대사적 질병들을 유발한다[1]. 근감소증(sarcopenia) 이라고 하는 이 질환과정은 특히 노인 환자에서 많이 발생하며, 건강과 삶의 질을 악화시킨다[2]. 근감소증은 평소에도 피로감을 높이고, 신체활동을 감소시키며, 여러 급성질환 등을 유도한다. 또한, 부상으로부터의 회복을 지연시키며, 만성질환으로 인한 사망률 증가와 입원기간의 연장 및 독립적인 생활을 하는데에도 많은 지장을 주는 것과 연관성이 높다[3-5].
골격근(skeletal muscle)은 우리 몸의 40%정도로 구성되어 있고, 운동과 대사의 균형적인 측면에서 매우 중요한 역할을 한다[6]. 또한, 근육은 우리 몸 안에서 가장 많은 단백질(protein)을 저장하는 기관이기도 하다[7]. 하지만, 근육 안의 단백질과 근육 내 단백질 대사의 감소는 근육량이 줄어드는 골격근 위축을 발생하게 한다[8]. 골격근 위축은 질병으로서 근감소증이라는 질병과 크게 연관되어 있고, 암[9], 심혈관계 질환[10], 유전적인 근육 질환[11], 신경학적 기능 장애[12] 등과 관련이 있다. 또한, 건강한 노인들에서도 골절(leg fractures)[13], 장시간의 비활동성, 척추 손상[14] 등과 관련이 있는 것으로 최근에 보고되었다. 이러한 골격근 위축의 초기 신호는 골격근 수축(muscle contraction)과 골격근 자극(stimuli)의 감소로 시작하여 근육 내 단백질 생성 대사 감소와 세포사멸사(apoptosis)촉진을 유도한다[15].
최근 세계보건기구(WHO)에서는 골격근 위축과 관련된 근감소증을 2017년에 질병 코드(ICD-10-CM)로 만들어 골격근 위축 관련 다양한 임상학적 연구를 진행하고 있다[16]. 이러한 시점을 기준으로 임상학적 접근 방법을 새롭게 개편하고 있고, 유럽의 최근 연구결과들과 가이드라인을 보면 근육의 신체적 기능과 골격근 양이 가장 중요한 변인 중 하나로 보고하였다[17]. 하지만, 광범위한 임상적 접근에도 불구하고, 분자생물학 연구분야에서 골격근 위축과 관련된 기전적 연구는 부족한 실정이다. 현재, 특별한 치료법이 필요한 상태이지만 의약학 분야에서는 골격근 위축 개선 약물 개발 등을 진행하고 있다.
한편, 많은 선행 연구들에서 비침습적인 방법으로 운동이 골격근 위축에 매우 긍정적인 역할을 한다고 보고하고 있다[18]. 하지만, 지금까지 골격근 위축에 대한 분자 수준에서 새로운 기전(mechanism)에 대한 설명은 부족한 실정에 있다. 실제로 몇몇 연구에서 밝혀진 기전은 영양학적 접근으로 골격근 위축에 대한 기전이 많은 실정이다. 이러한 영양학적 접근에서는 단백질 또는 아미노산(amino acids)의 섭취권장, 항산화제(antioxidant)와 같은 영양 보충을 중점적으로 설명하고 있다. 하지만, 운동과 영양은 서로 독립적이면서도 복합적인 연관성이 있음으로 본 연구에서는 운동을 중심으로 하는 골격근 위축의 기전적인 접근을 자세히 분석하고자 한다.
1. MurF1 & MAFbx
세포 수준에서 주된 골격근 위축 인자는 E3 ubiquitin ligases인 muscle RING finger 1 (MuRF1)과 muscle atrophy F-box, AKA atrogin-1 (MAFbx) 등이 있다. 이 인자들은 세포 수준에서 노화(aging), 당질 코르티코이드(glucocorticoid)의 증가, 근육 사용 감소 등의 다양한 스트레스(stress)환경에서 골격근 위축기전으로 작용된다[19]. 예를 들어, 이러한 골격근 위축과 관련한 세포수준의 기전은 당질 코르티코이드 수용기(receptor)와 Foxo 전사 인자(transcription factor)가 MuRF1의 유전자와 MAFbx의 단백질 발현을 증가시켜, 골격근 위축을 유발하게 된다[20]. 이와는 반대로 건강한 성인들에서는 Insulin-Like Growth Factor-1 (insulin/IGF-1) signaling을 통한 낮은 당질 코르티코이드 수치는 MuRF1, MAFbx의 발현과 Forkhead Box O Transcription Factors (FoxOs) 전사인자들의 감소를 유도하여 골격근 위축을 예방하는 것으로 나타난다[20]. 이러한 동화작용(anabolic action)의 중요한 역할을 하는 Insulin/IGF-1 signaling은 Akt/PKB (protein kinase B)의 직접적 FoxO 전사인자의 억제와 mechanistic target of rapamycin complex 1 (mTORC1)을 증가시켜 근육 내 단백질 대사 및 세포성장을 유도하게 된다[21].
2. IGF-1/Akt/mTOR/FoxOs
IGF-1과 인슐린(insulin) 작용으로 phosphatidylinositol-3-kinase (PI3K)/Akt 경로를 통해 근육의 크기는 조절이 된다[22]. 특히 이러한 PI3K/Akt가 활성화되어 mTOR가 자극을 받으면 70-kDa ribosomal S6 protein kinase (S6K), 4E-binding protein 1 (4E-BP1)가 인산화 작용(phosphorylate)을 통해 단백질 대사가 촉진하게 된다[23,24]. 따라서 Akt/protein kinase B는 골격근 위축의 세포대사, 세포성장 그리고 생존에 중요한 역할을 한다고 볼 수 있다. 인체 연구 분석을 보면, 12주간의 저항성 운동이 근육 무게의 변화를 크게 일으키는 것으로 나타났다[25]. 또한, 노인들에서 지속적인 저항성 운동을 통한 IGF-1의 발현과 근육의 증가는 큰 관련이 있는 것으로 보였다[26]. 하지만, 이러한 운동을 통한 정확한 기전에 대한 설명은 부족한 것으로 판단된다.
FoxOs집단은 근육 내 FoxO1부터 3으로 구성되어져 있고[27], FoxO라고 불리는 DNA 결합 부분의 영역을 방어하는 영역으로서 세포 핵에 위치하여 다양한 하부 단백질 신호전달(downstream signaling proteins)을 조절하는 전사인자로 알려져 있다[28]. 즉, Akt 신호 전달로 세포질 액상(cytosol) 내 FoxO가 인산화되는 과정을 통해 골격근 위축과 관련된 유전자(genes)들을 변형하지 못하게 한다[29]. FoxO 중 대표적인 FoxO1인자는 근육 내 동화작용 기전(anabolic pathway)의 효율을 높이고, 4E-BP1 단백질 전사 생성 물질의 인산화를 줄이는 것으로 보였다[28]. 이는 mTOR와 RAPTOR (regulatory-associated protein of mammalian target of rapamycin)의 감소로 나타났다. 또한, 인체 연구에서는 노인이(65세 이상) FoxO1의 근 핵(myonuclear) 수치가 젊은 사람들에 비해 높은 것으로 보였다[30].
3. Activating Transcription Factor 4 & Growth Arrest and DNA damage Inducible 45 Alpha (Gadd45a) & Cyclindependent Kinase 1a (P21/cdkn1a)
본 연구에서는 근육 위축이 이종 이량체 염기성 루신 지퍼(heterodimeric basic leucine zipper, bZIP) 전사인자의 하위인자인 전사인자 4 활성화 (ATF4: activating transcription factor 4)와 연관되어져 나타났다[31,32]. ATF4는 세포수준에서 항 동화작용(anti-anabolic cellular)에 유도하고, 이는 하위인자인 eIF2alpha kinase를 조절한다[31,32].
ATF4는 mTORC1의 하부 매개인자로서의 insulin/IGF-I 신호전달에 대한 동화작용을 유도한다[33]. 골격근에서 ATF4의 효과는 잘 알려져 있지 않으나, 근육 위축을 유발하는 기아(starvation), 암(cancer), 신부전(renal failure) 등에서 근육 위축과 ATF4 mRNA의 증가 사이의 연관성을 보였고, ATF4 mRNA의 수준의 증가는 MuRF1, MAFbx 및 다른 골격근 위축 관련 mRNA의 증가와 함께 발생했다[20,34]. 한편, 젊은 쥐의 근육 섬유에서의 ATF4 발현 억제는 금식과 limb immobilization을 통한 골격근 위축에 부분적으로 예방하는 것으로 보였다[35,36]. 따라서 ATF4는 노화, limb mobilization, 금식으로 인한 골격근 위축에 크게 관여하는 것으로 나타났다.
앞서 이야기한 ATF4의 역할과 기능에서 새롭게 추가된 인자인 Gadd45a와 p21은 직접적으로ATF4에 영향을 준다. 이러한 두 요소들은 골격근 위축으로 인해 근육 내 많이 발생되는 것으로 보고되어졌다[35-39]. 아직까지는 Gadd45a와 p21에 대한 정확한 기전은 밝혀지지 않았지만, 두 개의 동물 연구에서 밝혀진 바로는 쥐의 근육에서 골격근 위축 상태에서 ATF4의 발현은 증가하였다[36,40]. 또한 ATF4 경로의 하부 매개인자로서의 역할로, Gadd45a 및 p21은 MuRF1 mRNA 또는 MAFbx mRNA의 발현을 증가시키지 않으면서 근육대사를 위축을 촉진시킨다[40,41]. 이러한 발견은 ATF4 경로가 MuRF1 또는 MAFbx의 상위인자와 관련이 없고, MuRF1 및 MAFbx와 동일한 선상에서 역할을 하거나, MuRF1 및 MAFbx의 하위인자로서 역할을 한다고 분석하고 있다.
4. Exercise as Potential Therapies in Skeletal Muscle Atrophy
앞서 제시한 골격근 위축과 관련된 인자들이 운동을 통해 긍정적으로 개선되는 것을 알 수 있었다. 특히, 운동은 노화에서 몸 전체의 움직임을 통해 신체기능 또는 신체 수행 능력을 개선하고 골격근 비대(hypertrophy)와 근력(muscle strength)을 개선을 시키는 것으로 알려져 있다[24,42]. 본 연구에서는 골격근 위축 개선에 따른 운동의 역할에 대해 Table 1에 간략하게 요약을 하였다.
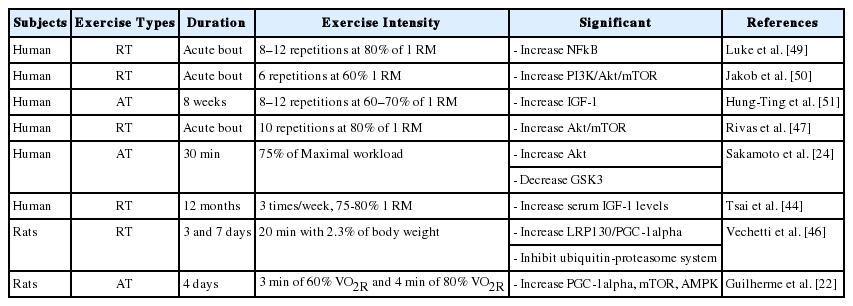
Summary of exercise-induced signaling pathway in skeletal muscle
운동은 일반적으로 유산소운동, 저항성 운동 및 복합운동으로 구분되는데, 본 연구에서는 운동의 종류를 고려하지 않고 운동 자체로서의 효과를 분석해 보았다. 이러한 운동은 골격근 위축 또는 근감소증에서 IGF-1/Akt/mTOR 신호 경로를 통해 골격근 위축을 예방하는 것으로 보였다. 특정 지역 내에 거주하는 노인들을 대상으로(65세 이상) 12주간의 저항성 운동을 통해 혈액 수준에서 IGF-1의 긍정적인 개선 확인하였고, IGF-1이 증가하면 제지방량(lean body mass)이 증가하는 것을 확인할 수 있었다[43]. 또한, 12개월 동안 저항성 운동을 시도하여 노인들의 IGF-1의 변화에 긍정적인 영향뿐만 아니라 homocysteine의 수치도 개선되는 것을 확인하였다[44]. 그리고 젊은 사람들과 비교했을 때, 노인들에서 IGF-1의 수치가 운동을 통해 훨씬 더 크게 개선되는 것이 확인되었다[45]. 그리고 동물 실험 연구에서는 노화 동물 쥐에서 유산소성 운동을 통해 FoxO 인자가 leucine rich protein 130 (LRP130)/peroxisome proliferator-activated receptor gamma-coactivator-1α (PGC-1 α)의 신호 자극으로 골격근 위축 억제하는 역할을 하는 것으로 나타났다[46]. 또한 이러한 노화 동물에서 저항성 운동을 통해 Nuclear factor-κB (NFkB)의 신호와 ceramide 구성 요소가 증가되는 것으로 보아 운동이 골격근 위축에 긍정적인 역할을 하는 것으로 보인다[47]. 특히, 인체연구에서는 여성 노인들에서 home-based 유산소성 운동이 NFkB와 관련 유전자 표현형에 영향을 주는 것으로 보였다[48].
결 론
골격근 위축증은 복잡한 분자 신호 전달을 통해 진행되며, 본 연구에서는 운동이 전반적으로 IGF-1/Akt/mTOR의 신호 전달을 활성화시키고, 골격근 위축 관련 인자들인 MurF1 & MAFbx, IGF-1 및 NFkB 등에 긍정적으로 많은 역할을 하는 것으로 분석하였다. 특히, 본 연구에서는 골격근 위축에 관여하는 새로운 인자로 ATF4, Gadd45a와 p21에 대해 설명을 하였지만, 이와 관련하는 운동의 효과를 검증한 연구결과는 전무한 것으로 보였다. 그러나 골격근 위축에 동반되는 기존에 밝혀진 인자들 뿐만 아니라 새로운 인자들을 함께 운동과 훈련을 통해 분석한다면, 더욱 명확한 골격근의 감소와 증대에 관한 분자생물학적 기전분석결과를 기대할 수 있을 것이다.
Notes
이 논문 작성에 있어서 어떠한 조직으로부터 재정을 포함한 일체의 지원을 받지 않았으며, 논문에 영향을 미칠 수 있는 어떠한 관계도 없음을 밝힌다
AUTHOR CONTRIBUTION
Conceptualization: DY Seo; HS Bang; Formal analysis: JH Bae, HS Bang; Methodology: DY Seo; Project administration: JH Bae, HS Bang; Writing-original draft: DY Seo, YS Kwak; Writing-review & editing: YS Kwak