High-Intensity Resistance Training Suppresses Exacerbation of Atopic Dermatitis in Mice
Article information

Abstract
Abstracts
Purpose
Training generally promotes health and inhibits diseases. However, in patients with atopic dermatitis (AD), training and sweating are significant aggravating factors. This study examined the effect of high-intensity resistance training (HIRT) on AD-like skin lesions in mice.
Methods
Eight-week-old female BALB/c mice were divided into four groups (control, HIRT, AD-only, and AD+HIRT). The mice in the HIRT group performed vertical ladder climbing for four weeks.
Results
After four weeks of HIRT, histopathological examination revealed reduced epidermis/dermis and dermal infiltration of inflammatory cells in the mice ear tissue. Additionally, HIRT suppressed serum immunoglobulin (Ig) levels and mRNA expression of pathogenic cytokines in the ear tissue; further, it reduced the size and weight of the draining lymph node (dLN) and non-dLN (ndLN), and the pathogenic cytokine-related mRNA expression of CD4+T cells from dLNs and ndLNs. We thus observed a negative correlation between HIRT and AD symptoms in mice.
Conclusions
The results show that HIRT exerts positive effects in patients with AD.
INTRODUCTION
Atopic dermatitis (AD) is a chronic inflammation of unidentified origin associated with dry skin, erythema, and eczema, and usually begins in early infancy but also affects a significant number of adults [1–3]. However, AD has been reported to be related to genetic and environmental factors such as allergens and microbes along with immune system and skin barrier dysfunction. It has also been reported that acute and chronic phase AD are strictly related to T helper 2 (Th2) and T helper 1 (Th1) cell-mediated AD lesions, respectively [3,4]. In addition, mast cell activation, elevated IgE levels, IgG and pro-inflammatory cytokines including interleukin (IL)-4, IL-13, IL-31, tumor necrosis factor-al-pha (TNFα), and interferon-gamma (INFγ) play a vital role in skin abrasions [5–9]. Accordingly, the role of IL-17 has been suggested in certain skin conditions [10,11].
Training is a well-known practice for maintaining good health and preventing diseases [12]. Regular training is effective in reducing the risk of coronary artery disease, stroke, hypertension, and osteoporosis [13]. Meanwhile, it is plausible that AD patients do not training because of the itching due to sweating during training. This hypothesis is supported by previous studies by Stern et al. [14] and Williams et al. [15]. On the contrary, there is other aspect that regular physical activity may be beneficial to patients with AD [16]. In addition, to the best of our knowledge, the effect of high-intensity resistance training (HIRT) on AD has not yet been evaluated. In this study, therefore, we evaluated the effect of HIRT on AD lesions using BALB/c mice.
METHODS
1. Animals
Eight-week-old female BALB/c mice (Samtako, Osan, Republic of Korea) were housed under specific pathogen-free conditions. Five to ten mice per cage were housed in a laminar airflow room, maintained at a temperature of 22±2°C with a relative humidity of 55±5% throughout the study. All experiments were approved by the Institutional Animal Care and Use Committee of Konkuk University.
2. Induction of AD lesions in the ear and high-intensity resistance training protocol
AD was induced in the mice by repeated local exposure of the ears to Dermatophagoides farinae extract (DFE; house dust mite extract) and 2,4-dinitrochlorobenzene (DNCB) as previously described [17]. For AD induction, the mice were divided into four groups (control, HIRT, AD-only, and AD+HIRT). The surfaces of both earlobes were stripped five times with surgical tape (Nichiban, Tokyo, Japan). After stripping, 20 µL of 1% DNCB was applied on each ear, followed 4 days later by 20 µL of DFE (10 mg/mL). DFE or DNCB treatment was administered alternately, once a week for 4 weeks.
The HIRT performed climbing of a vertical ladder for 4 weeks. The training was performed using a 1-m ladder with a 1.5-cm grid and inclined at 85° (Fig. 1A). Initially, the mice performed climbing with free weight for a week in order to get accustomed to the training. For the first training session, 10% of their body weight was attached to the base of their tails during a week of training, and the resistance was progressively increased to 0% (2 times), 10% (2 times), 30% (2 times), and 50% (2 times) during the next 2 weeks. For the last training session, 100% of their body weight was attached to the base of their tails during a week.

(A) Ladder for high-intensity resistance training (HIRT). (B) Experimental schedule for the induction of AD lesion. (C) The mice performed climbing of the vertical ladder for 4 weeks. The training was accomplished using a 1-m ladder with a 1.5-cm grid and inclined at 85°. Initially, the mice performed climbing with free weight for a week in order to become accustomed to the weight. For the first training session, a free weight, which was 10% of the body weight, was attached to the base of their tails, and the resistance was progressively increased to 30% and 50% during the 4 weeks.
3. Ear thickness measurement and blood samples preparation
Ear thickness was measured 24 hours after DNCB or DFE application with a dial thickness gauge (Kori Seiki MFG, Co., Japan). At days 14 and 28, blood samples were collected via orbital puncture. Plasma samples were prepared from the blood samples and stored at −70°C for further analysis. After blood collection, the ears were removed and used for histopathological analysis. Serum immunoglobulin (Ig) E and IgG2a levels were measured at days 14 and 28 after the first induction using an IgE enzyme-linked immunoassay kit (Bethyl Laboratories, Inc., Montgom-ery, TX, USA) according to the manufacturer’ s instructions.
4. Histological observations
The excised ears were fixed in 4% paraformaldehyde for 16 hours and embedded in paraffin. Thin (6 µm) sections were stained with hematoxylin and eosin (H&E). The thickness of the epidermis and dermis was measured under a microscope. For measurement of mast cell infiltration, skin sections were stained with toluidine blue, after which the number of mast cells was counted in five randomly chosen fields of view.
5. Real-time polymerase chain reaction (PCR)
Quantitative real-time PCR was carried out using a Thermal Cycler Dice TP850 (Takarabio Inc., Shiga, Japan) according to the manufacturer’ s protocol. Total RNA was isolated from the ear tissue and lymph nodes of each group. The PCR conditions were similar to those previously described [17]. Two microliters of cDNA (100 ng), 1 μL of sense and antisense primer solution (0.4 µM), 12.5 μL of SYBR Premix Ex Taq (Takarabio Inc.), and 9.5 μL of dH2 O were mixed to obtain a final 25-μL reaction mixture in each reaction tube. The amplification conditions were as follows: 10 seconds at 95°C, 40 cycles of 5 seconds at 95°C and 30 seconds at 60°C, 15 seconds at 95°C, 30 seconds at 60°C, and 15 seconds at 95°C. The mRNA levels of the target genes relative to glyceralde-hyde 3-phosphate dehydrogenase (GAPDH) were normalized using the following formula: relative mRNA expression=2-(Ctoftarget gene−Ct of GAPDH), where Ct is the threshold cycle value. In each sample, the expression level of the analyzed gene was normalized to that of GAPDH and presented as a relative mRNA level.
6. Statistical analysis
Statistical analysis was carried out using SAS statistical software (SAS Institute, Cary, NC, USA). Multiple group data were analyzed using one-way analysis of variance followed by Dunnett’ s multiple range test. All results are expressed as the mean±standard deviation of comparative fold differences. Data are representative of three independent experiments. The threshold for significance was set at p <.05.
RESULTS
After HIRT for 4 weeks, we observed that AD+ HIRT significantly reduced the thickness of both the right and left ears (Fig. 2A, B) (p <.05). Also, a reduced number of infiltrated mast cells was found in the AD+HIRT group mice, compared with the AD group (Fig. 3A, C, and D). Furthermore, AD+HIRT suppressed the thickness of epidermal and dermal cells (Fig. 3B). We found a reduced number of mast cells in the AD+HIRT group mice (Fig. 3B).

Photographs of the ears of mice from each group on day 28. (A) Right ear (B) left ear thickness was measured with a dial thickness gauge every 3 days after 2,4-dinitrochlorobenzene (DNCB) or Dermatophagoides farinae extract (DFE) application. *indicates a significant difference from the value of the DFE/DNCB-treated mice at p<.05. AD, atopic dermatitis; HIRT, high-intensity resistance training.
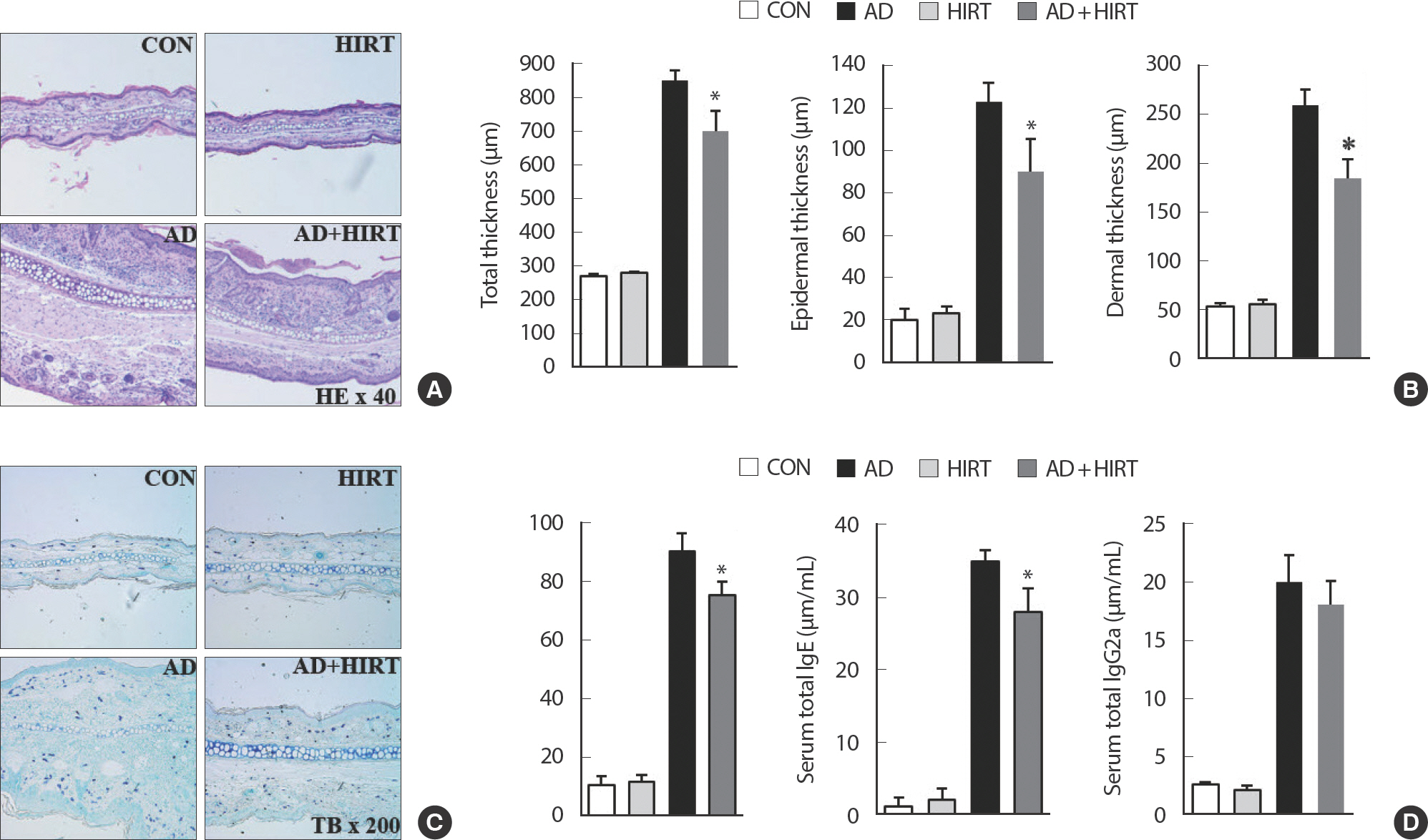
Representative photomicrographs of ear sections stained with hematoxylin and eosin (A) or toluidine blue (B). Epidermal and dermal thickness were measured using microphotographs of hematoxylin and eosin-stained-tissue (C). The number of infiltrated mast cells was determined based on toluidine blue staining (D). Blood samples were collected via orbital puncture at day 28. Plasma IgE (E) and IgG2a (F) levels were quantified using enzyme-linked immunosorbent assay. Data are presented as the mean±SD of triplicate determinations. *indicates a significant difference from the value of the DFE/DNCB-treated mice at p<.05. AD induced by DFE and DNCB treatment. The pictures shown are representative of each group (n = 3–6). The original magnification was 100×CON. AD, atopic dermatitis; HIRT, high-intensity resistance training.
AD+HIRT significantly suppressed the expression of Th2-related cytokines such as IL-4, IL-6, IL-10, IL-13, and IL-31 as well as Th1-related cytokine such as IFN-γ in the ear tissue (Fig. 4). We evaluated the size and weight of dLN and ndLN as well as the cytokine-related mRNA expression of CD4+T cells from dLN and ndLN cells. The AD mice had bigger dLN compared with the healthy untreated control mice (Fig. 5A). The size of dLN were reduced after 4 weeks of HIRT (Fig. 5A). However, no significant changes were observed among the four groups of mice regarding their ndLN size (Fig. 5B). Investigation of the inflammatory mRNA-related cytokines from CD4+T cells purified from dLN revealed that the mice in the AD+HIRT group had a significantly lower expression of IL-4, L-17, and I INFγ (Fig. 5A). However, the AD+HIRT group mice did not affect mRNA expression of IL-4, L-17, and I INFγ in the CD4+T cells purified from their ndLNs (Fig. 5B).
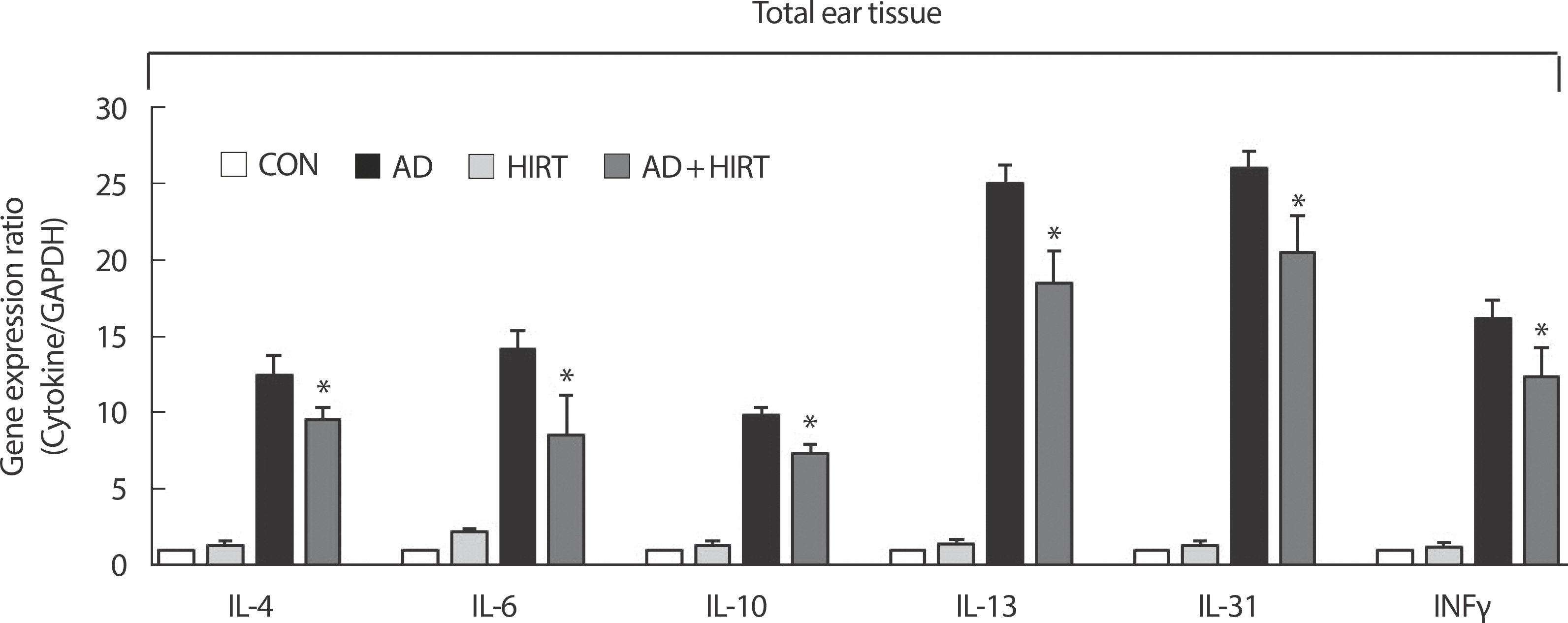
Effect of high-intensity resistance training (HIRT) on the expression of various pathogenic factors in the ear. The ears were excised on day 28 and total RNA was isolated. Quantitative real-time polymerase chain reaction was performed as described in the Methods section. Data is presented as the mean±SD of triplicate determinations. *indicates a significant difference from the value of the DFE/DNCB-treated mice at p<.05. AD induced by DFE and DNCB treatment. DNCB, 2,4-dinitrochlorobenzene; DFE, Dermatophagoides farina extract; CON, control; AD, atopic dermatitis.
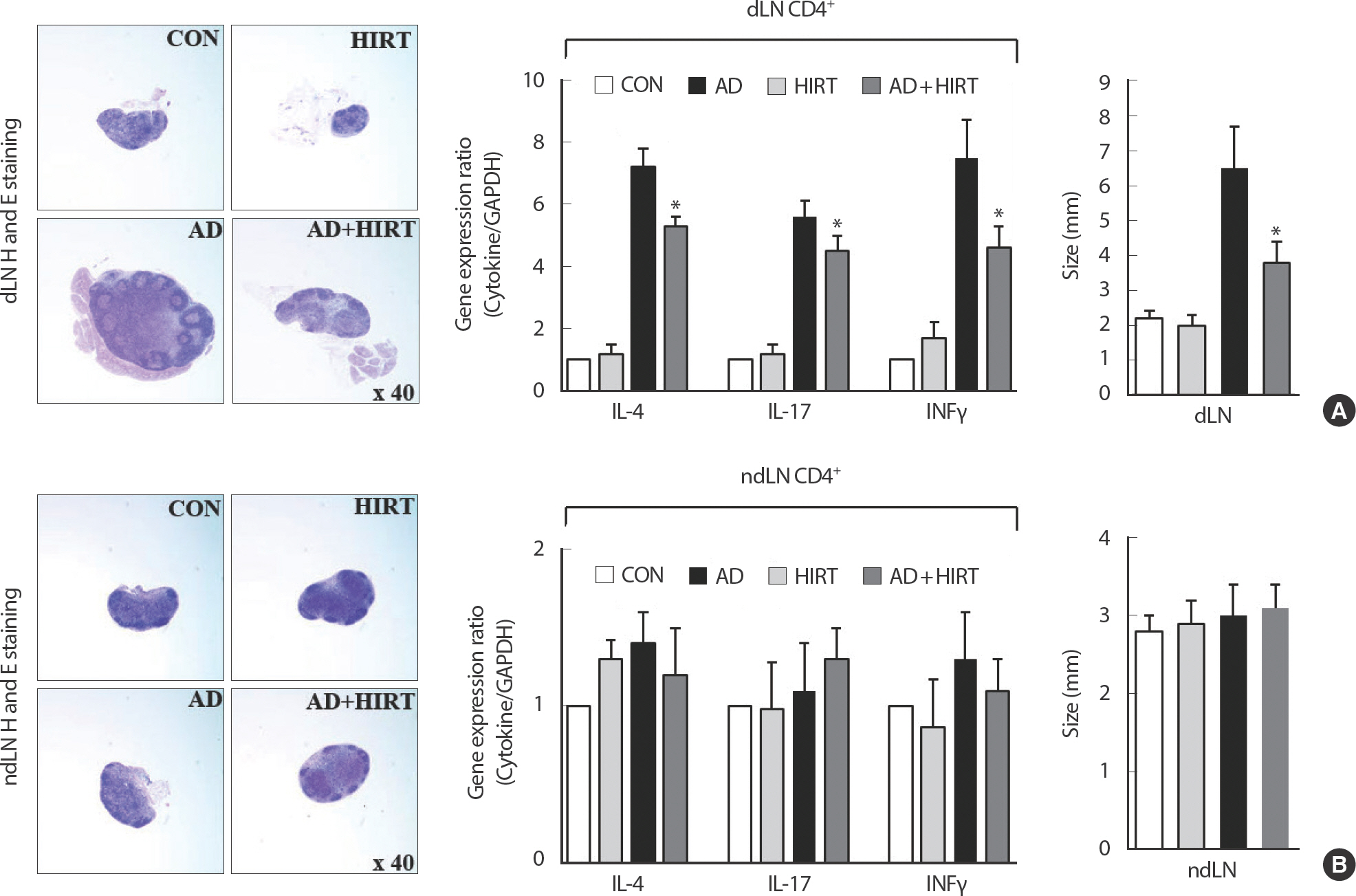
Effect of high-intensity resistance training (HIRT) on the expression of size of draining lymph node (dLN) and non-dLN (ndLN), and the cytokine-related mRNA expression of CD4+T cells from the dLN (A) and ndLN (B). *indicates a significant difference from the value of the DFE/DNCB-treated mice at p<.05. AD induced by DFE and DNCB treatment. DNCB, 2,4-dinitrochlorobenzene; DFE, Dermatophagoides farina extract; CON, control; AD, atopic dermatitis.
DISCUSSION
Training is necessary for a healthy life. Regular training not only improved antioxidant activities and immunity but also reduced the danger of some diseases [13,18,19]. However, Williams et al. [15] and Stern et al. [14] reported that training that cause sweating were notable AD worsening factors in schoolchildren. Accordingly, researchers have also reported that training, heat, and sweating, caused the most discomfort for school-children with AD in Singapore [20]. Nevertheless these previous studies, there were little investigation systemic analysis of the effect of high intensity training on AD. Afterward, we hypothesize that training may have positive effect on AD patients. Thus, in the present study, we evaluated the effect of HIRT on rat models with AD. To examine the immu-nomodulatory properties of HIRT, the mice performed climbing of a vertical ladder for 4 weeks. After HIRT for 4 weeks, we observed ear thickness, inflammatory cell infiltration, IgE and IgG2a expression in serum, mRNA level of immunoregulatory cytokines in the ear tissue and the size, weight of dLN and ndLN, and pathogenic cytokine-related mRNA expression of CD4+T cells from dLNs and ndLNs. The cross-linking of IgE and allergen stimulates the mast cells that release cytokines at the sites of allergic reactions [21]. Further, increased levels of IgG antibodies have been reported to play a role in AD and have been associated with Th1 response [22]. A number of studies have reported that AD shows increased total IgE level and Th2/Th1-type cytokine expression [23–26]. To define whether AD exerts its effects mainly via Th1 or Th2 response, serum levels of IgE (total and DFE-specific) and IgG2a were measured in each treatment group and reduced total IgE and IgG2a levels were observed in AD+HIRT group mice, compared with those of the DFE/DNCB-treated mice. These results propose that the effect HIRT on AD development is related with the downregulation of serum Ig levels.
When pathogens are encountered, naive CD4+T cells lead to the generation of Th1 and Th2 cells. Th2 cells produce IL-4, IL-13, and IL-17, which encourage humoral immunity and produces IgE as well as control Th1 response [27]. In particular, IL-4 plays an important role in the transformation of naive CD4+T cells into Th2 cells [28]. Furthermore, AD has been reported as a systemic disease [29]. Therefore, we further examined whether HIRT affects systemic immune responses. In a previous study using NC/Nga mice, it was revealed that AD-like symptoms were promoted by strong training and reduced by mild training [30]. Accordingly, Kim et al. [17]. reported that AD symptoms increased with high-intensity swimming training in BALB/c mice with higher IgE levels and cytokines expression. Differing to these investigations, our study shows that HIRT suppressed AD symptoms.
CONCLUSIONS
We found that the severity of AD evidenced by histopathological signs, IgE, IgG levels, and expression of mRNA of pathogenic cytokines in the ear tissue was suppressed by HIRT. In addition, HIRT reduced the size of dLN, and pathogenic cytokine-related mRNA expression of CD4+T cells from dLNs, whereas the weight and size of ndLN were not significantly different. This results suggests that HIRT could be beneficial for people with AD.
Acknowledgements
This work was supported by research grants from Daegu Catholic University in 2018.
Notes
The authors declare no conflict of interest.
AUTHOR CONTRIBUTION
Conceptualization: E Choi, J Kim; Data curation: KC Cho; Formal E Choi, KC Cho; Methodology: E Choi, KC Cho; Visualization: KC Cho; Writing-original draft: E Choi, J Kim.