서 론
Irisin은 운동 유발성 마이오카인(exercise-induced myokine)으로 Boström et al. [1]의 연구에 의하면, 생쥐의 골격근에서 막 단백질인 fibronectin type III domain-containing protein 5 (FNDC5)에서 분해되어 혈액으로 방출된다고 보고되고 있다. FNDC5는 운동에 반응하여 peroxisome proliferator-activated receptor-γ coactivator-1α (PGC1α)에 의해 조절되는 것으로 알려져 있고, PGC1α는 운동 후 골격근에서 발현되며 체중 감소와 산화 스트레스 조절 기전에 관여할 뿐만 아니라 인슐린 신호 전달 경로의 효율성을 향상시켜 인슐린 민감성을 향상시키는 것으로 알려져 있다[2-4].
이 보조 인자를 매개하는 방법을 밝히기 위해 Boström et al. [1]은 3주 동안 자발적 휠 러닝(wheel running) 운동 프로그램을 시행한 생쥐를 이용하여 골격근에서 PGC1α의 과발현(overexpression)이 피하지방(subcutaneous fat)의 uncoupling protein 1 (UCP1) 상향 조절에 관련이 있음을 관찰했다. 즉, PGC1α의 과발현으로 인해 지방 세포의 갈변(browning)이 유도되는 것을 발견했으며, 이때 잠재적으로 관여하는 인자들 중 하나인 FNDC5가 지방 조직의 갈변을 매개할 수 있는 단백질로 제시되었다[1]. 더욱이 이 단백질이 어떻게 절단되어 irisin 형태로 혈액 순환에 방출되는지를 보여 주었고 이후 irisin은 운동 유발성 호르몬으로 가정되었다[1].
Irisin은 미토콘드리아 내막에 존재하는 UCP1 수준을 증가시켜 백색지방 세포의 갈변을 유도하고 열 생성을 유도하는 것으로 제안되었으며, 생쥐 모델에서 irisin은 총 신체 에너지 소비를 증가시키고 비만으로 유발되는 인슐린 저항성에 긍정적인 역할을 하는 것으로 알려져 있다[1,5]. 생쥐와 마찬가지로 FNDC5 유전자는 인간의 근육세포에서도 발현되는 것으로 나타났으며[6], 인간의 지방 조직에서 irisin에 의해 유도된 베이지 지방 세포(beige adipocyte)의 자극은 비만과 관련된 대사 장애를 개선하는 데 활용 가능한 치료 방법으로 제시되고 있다[7]. 백색지방에서 발견되는 베이지 지방 세포는 체중 감소 및 글루코스 대사에 긍정적인 영향을 미치며, UCP1의 발현을 증가시킨다고 알려져 있다[8,9]. 그뿐만 아니라, 또 다른 선행연구들에 의하면 irisin은 심혈관 질환, 제2형 당뇨병 또는 지방간과 같은 질환에 대해 보호 역할을 할 수 있으며, 비만 및 골다공증, 신경 퇴행성 질환뿐만 아니라 비만으로 유발되는 암의 예방에도 잠재적인 역할을 할 수 있는 것으로 알려져 irisin이 대사성 질환의 잠재적인 치료 표적으로 주목받고 있다[10-12].
Irisin이 운동 유발성 호르몬으로 알려져 있음에도 불구하고 최근 연구들에 의하면 정기적이고 규칙적인 유산소 운동이 혈중 irisin의 농도를 증가시키지 못했다는 연구들도 보고되고 있다. Hecksteden et al. [13]의 연구에서, 건강한 성인을 대상으로 26주 동안 주 3회 45분 중강도 유산소 트레이닝을 실시한 결과 혈청 irisin이 유의하게 증가하지 않았으며, Pekkala et al. [14]의 연구에서도 건강한 중년 남성을 대상으로 21주 동안 고강도 유산소 운동을 실시한 후에 혈청 irisin의 농도가 증가하지 않았다고 보고하였다.
반면, Norheim et al. [3]의 연구에서는 1회성(acute) 및 규칙적인 장기(chronic) 운동 후 나타난 혈장 irisin의 변화를 살펴본 결과, 운동 전과 비교하여 irisin의 농도가 1회성 유산소 운동 후 급격하게 증가하였으며, 규칙적인 12주간 장기 유산소 운동 후에는 오히려 감소한 결과를 나타냈다. 또한, Huh et al. [6]의 연구에서는 8주간의 중강도 유산소 트레이닝이 irisin 수준에 아무런 영향을 미치지 않았음을 나타냈으며, 오히려 1주 동안 중강도 유산소 트레이닝 후 혈중 irisin 수준이 증가한 결과를 제시하였다[6].
이렇듯 irisin은 운동 직후에 증가하고 운동 강도와 상호 관련이 있다고 선행연구들에 의해 제시되고 있다[6]. Tsuchiya et al. [15]의 연구에서는 건강한 성인 남성을 대상으로 유산소 운동을 실시한 결과, 혈청 irisin이 저강도 유산소 운동(VO2max의 40%)보다 고강도 유산소 운동(VO2max의 80%) 후에 더 큰 증가 반응을 보였으며, Huh et al. [16]의 연구에서도 건강한 청소년을 대상으로 수영 운동을 실시한 결과, 지속적 중강도 수영 운동보다 고강도 인터벌 수영 운동이 보다 높은 혈중 irisin 수준을 나타내었다. 하지만, 이와 대조적으로 Winn et al. [17]의 연구에서는 비만 여성을 대상으로 중강도 유산소 운동(VO2max의 55%)과 고강도 인터벌 유산소 운동을 실시한 결과, 중강도 운동을 실시했을 때 혈장 irisin 농도 상승이 오랫동안 지속되었다. 선행연구들을 종합해 본 결과, 유산소 운동 강도에 따른 irisin 발현량의 차이에 대한 명확한 결과가 부족하여 추가연구가 필요한 실정이다. 또한 irisin이 운동호르몬으로 가정되었다는 사실에도 불구하고, irisin 발현에 있어 운동의 영향에 대한 기전은 명확하지 않다.
따라서 본 연구에서는 건강한 수컷 와일드 타입(wild type) 생쥐를 대상으로 운동 강도에 따른 irisin의 발현 정도의 차이를 비교하고자 하며, irisin의 상향 조절인자로 알려진 PGC-1α, FNDC5 그리고 irisin의 목표 조직인 지방의 UCP1 발현량 차이를 살펴봄으로써 트레드밀 유산소 운동의 효과에 대한 irisin 관련 기전을 규명하고자 한다.
연구 방법
1. 연구 대상
이 연구는 생후 13-15주 된 건강한 수컷 와일드 타입(wild type) C57BLKS/J-m+/m+ 생쥐를 대상으로 이루어졌으며, 4그룹으로 생쥐를 각 5마리씩 무작위로 배정하였다. 동물을 일정한 조명(12:12h light/dark), 상대습도(50-80%), 실내온도(20 ±1°C)의 환경 제어 조건하에서 사육하였으며, 물과 사료는 자유롭게 제공되었다. 모든 실험은 인천대학교의 동물 보호위원회의 승인(인천대 동물실험 2017-11)을 받은 후 수행하였다.
2. 연구절차
1) 운동 처치
통제 그룹을 제외한 나머지 실험용 생쥐는 설치류를 대상으로 제작된 트레드밀 JD-A-09 (JEUNG DO BIO & PLANT CO., Ltd, Korea)를 이용하여 저강도(low-intensity exercise, LIE), 중강도(moderate-intensity exercise, MIE), 고강도(high-intensity exercise, HIE) 운동을 실시하였다. 생쥐는 본 운동을 실시하기 전 준비운동(10 m/min, 경사도 0°, 5분)을 하였으며, 이후 운동 강도에 따라 저강도(10 m/min, 경사도 0°, 25분), 중강도(15 m/min, 경사도 0°, 25분), 고강도(20 m/min, 경사도 0°, 25분)로 나누어 운동을 실시하였다. 운동 강도는 생쥐를 대상으로 한 선행연구들을 참고하여 설정하였으며[18-22], 모든 운동은 1회성으로 진행하였다. 한 번의 운동으로 운동 직후 irisin의 반응과 irisin과 관련된 신호전달 단백질의 급성 반응(acute response)을 살펴보았다.
2) 젖산
유산소 트레드밀 운동이 적절히 처치되었는지를 확인하기 위해 실험용 생쥐의 꼬리를 잘라 전혈(약 32 μL)을 추출하여 accutrend plus (Mannheim, Germany)를 사용하여 운동 전과 후에 젖산을 측정하였다.
3) 혈당
혈당의 측정은 4그룹 모두 운동 전 생쥐의 꼬리를 잘라 전혈(약 10 μL)을 추출하여 혈당검사지(test strip)에 혈액을 묻혀 accu-chek performa (Roche, Korea)를 사용하여 측정하였다.
4) 몸무게
몸무게의 측정은 4그룹의 생쥐 모두 2.5% tribromoethanol (0.01 mL/g of body weight)을 복강(I.P)에 주사하여 마취시킨 후 전자저울(mettler toledo, Switzerland)을 사용하여 측정하였다.
5) 조직 적출
생쥐를 마취시켜 몸무게를 측정하고 복강 내의 대정맥(vena cava)으로부터 혈액을 추출하였다. 이후 생쥐의 내장(visceral) 부분에서 백색지방(white adipose tissue)을 추출한 후 양쪽 다리에서 비복근(gastrocnemius)과 가자미근(soleus)을 추출하였으며, 마지막으로 갈색지방(brown adipose tissue)을 견갑골 사이(interscapular) 부분에서 추출하였다. 적출된 조직들은 액화질소를 이용하여 급속 냉동시켰으며 분석 전까지 -80°C에서 보관하였다.
6) Enzyme-Linked Immunosorbent Assay (ELISA) 분석
Irisin 분석을 위해 마취된 생쥐의 복강 내의 대정맥으로부터 전혈을 추출하여 혈액응고를 위해 실온(room temperature)에 30분간 보관하였다. 이후 10분간 3,000 rpm으로 원심 분리한 후 혈청(serum) 샘플을 채취하였으며, 분석 전까지 -80°C에서 보관하였다. 혈청 irisin 농도는 상업적으로 이용 가능한 ELISA kit (Phoenix Pharmaceuticals, CA, USA, EK-067-29)를 사용하여 제작회사의 실험 절차에 따라 측정하였다.
7) Western blot
가자미근과 갈색지방의 단백질 분석은 전기영동법(western blot)에 의해 분석하였다. 이 실험에서는 Bio-rad사의 western blot 시스템을 사용하여 전형적인 형태의 전기영동법을 사용하여 특정한 단백질의 양을 분석하였다. 채취된 가자미근과 갈색지방은 protease inhibitors cocktail (Sigma-aldrich)이 섞인 CelLytic MT lysis buffer (Sigma-Aldrich)로 균질화시켰다. Bicinchoninic acid (BCA) protein assay를 통해 전체 단백질 양을 측정한 뒤, sodium dodecyl sulphate (SDS) -polyacrylamide gel을 이용한 전기영동을 실시하고 nitrocellulose membrane으로 전이시킨 후 5%의 탈지우유(skim milk)로 차단(blocking)시켰다. 이후, β-actin (Santa Cruz Biotechnology, 1:1,000)은 밤새(over-night) 4°C에서 배양(incubation) 시켰으며, FNDC5 (abcam, 1:500)와 UCP1 (abcam, 1:1,000) 역시 밤새 4°C에서 배양시켰다. 배양 후 tris-buffered saline (TBS)과 Tween 20을 섞은 TBST를 사용하여 wash를 5분간 2번, 10분간 2번 총 30분 실시하였으며 β-actin은 Chemidoc Touch Imaging System (Biorad, USA)을 사용하여 최종 밴드 강도를 정량화하였다. FNDC5와 UCP1은 secondary 항체(abcam, 1:2,000)와 함께 실온에서 1시간 배양하였다. 이후 앞서 설명한 방법으로 wash를 5분간 2번, 10분간 2번 총 30분 실시한 후 Chemidoc Touch Imaging System (Biorad, USA)을 사용하여 최종 밴드 강도를 정량화하였다.
8) 중합효소연쇄반응법(Quantitative Real-Time PCR, q-PCR)
전체 가자미근, 비복근, 백색지방 그리고 갈색지방은 cDNA (maxime RT PreMix kit (Intron Biotechnology, Seoul, Korea)로 역전사되었다. cDNA의 Real-time PCR 증폭은 Bio-Rad CFX-96 real-time PCR system (Bio-Rad, Hercules, CA)을 사용하여, SYBR Green Realtime PCR Master Mix (Toyobo, Japan)로 수행하였다. 데이터는 β-actin으로 일반화하였다. Primer sequences는 Table 1에 제시하였다.
3. 자료처리방법
모든 데이터는 평균 ±표준 오차(SEM)로 표시하였다. 모든 통계 분석은 GraphPad Prism6.05 version (La Jolla, CA 92037 USA)을 사용하여 분석하였다. 운동 전후에 따른 젖산은 paired t-test를 사용하여 분석되었으며, 운동 강도에 따른 irisin과 PGC1α, FNDC5, UCP1의 발현 정도는 일원 배치 분산분석(One-way ANOVA)을 통해 분석하였고, 사후 검증은 Tukey’s multiple comparisons test를 이용하여 분석하였다. 모든 통계적 유의 수준은 p<.05로 하였다.
연구 결과
1. 생쥐의 기초적 특성
생쥐의 몸무게, 글루코스, 운동 전후 젖산과 같은 기초적인 특성은 Table 2에 제시하였다. 모든 그룹 간 몸무게, 혈당, 운동 전 젖산, 운동 후 젖산은 유의한 차이가 나타나지 않았으며, 1회성 유산소 운동 강도에 따른 생쥐의 운동 전과 후의 혈중 젖산 농도는 모든 그룹에서 유의한 차이가 나타나지 않았다.
3. 운동 강도에 따른 FNDC5 발현
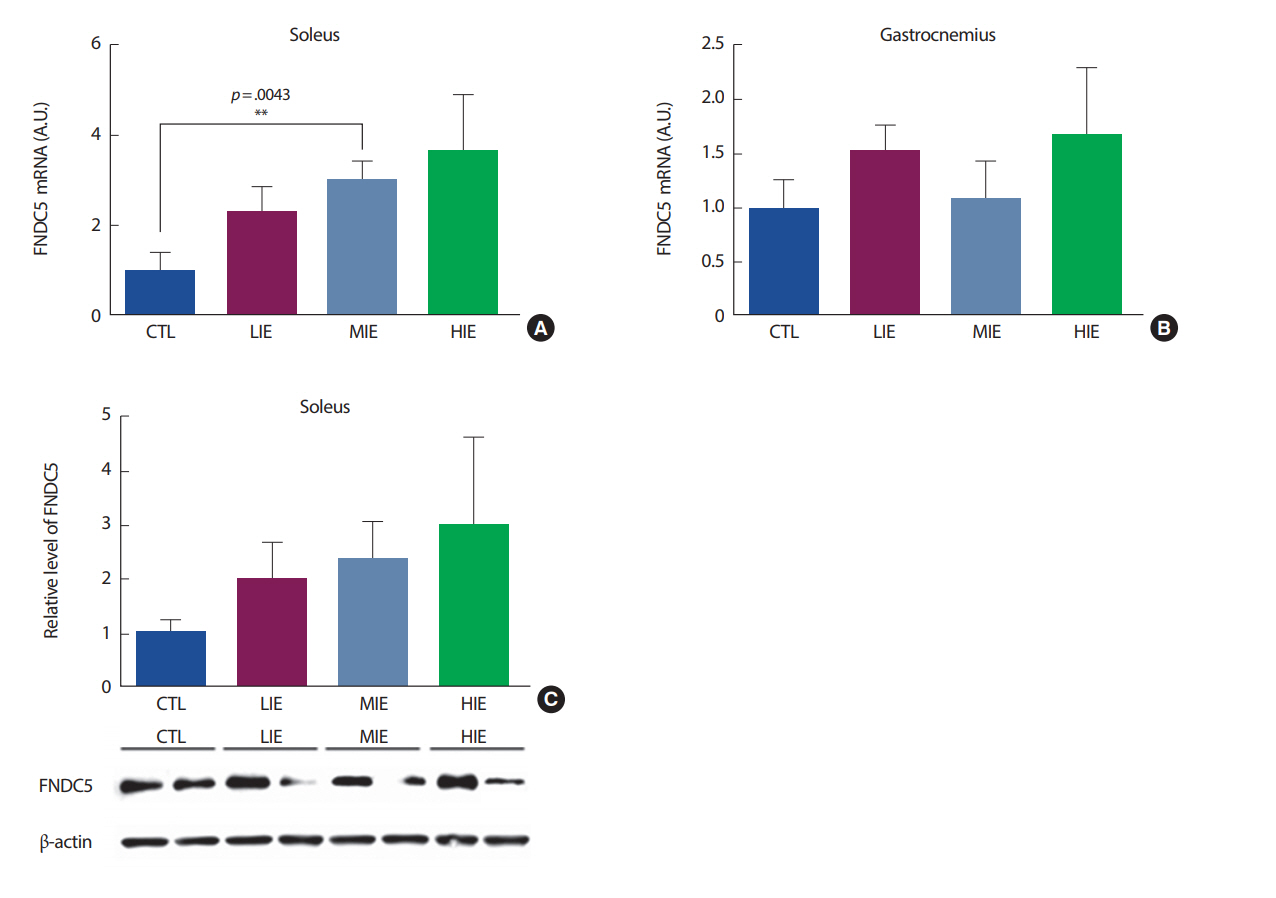
q-PCR 방법을 사용하여 1회성 유산소 운동 후 가자미근과 비복근의 FNDC5 발현 수준을 살펴보았다. 전형적인 Type I 근육으로 알려져 있는 가자미근의 경우, 운동 강도가 증가할수록 FNDC5 mRNA의 발현이 증가하는 경향이 나타났으며, 통제(control, CTL) 그룹과 중강도 운동 그룹(MIE) 간에 유의한 차이(p =.0043)가 나타났다(Fig. 2A). 이와는 달리 비복근의 경우, FNDC5 mRNA 발현에 그룹 간 유의한 차이가 나타나지 않았다(Fig. 2B). 추가적으로 FNDC5의 단백질 발현 정도를 살펴보기 위해 western blot을 실시한 결과, 운동 강도가 증가할수록 증가하는 경향을 보였으나 통계적으로 유의한 차이가 나타나지 않았다(Fig. 2C).
5. 운동 강도에 따른 혈청 irisin 농도
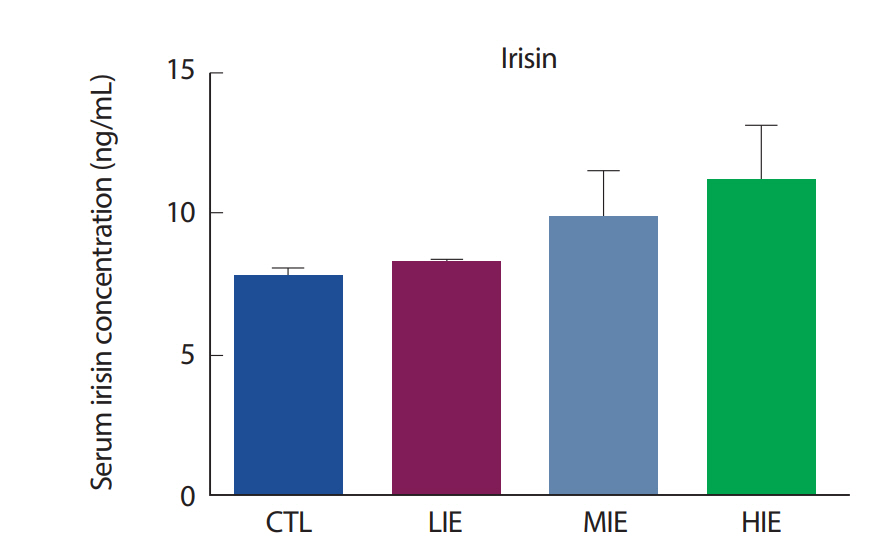
상업적으로 이용 가능한 irisin ELISA kit (Phoenix Pharmaceuticals, CA, USA)를 사용하여 유산소 운동 강도에 따른 혈청 irisin의 농도를 살펴본 결과, 중강도 및 고강도 유산소 운동 후 증가하는 경향을 나타냈지만, 모든 그룹 간 통계적으로 유의한 차이는 나타나지 않았다(Fig. 5).
논 의
이 연구에서는 생후 13-15주 사이의 건강한 수컷 wild type C57BLKS/J-m+/m+ 생쥐를 대상으로 1회성 유산소 운동 강도에 따른 irisin 및 FNDC5 발현 정도의 차이를 비교하고자 하였으며, 운동 직후 이 단백질들의 발현에 가장 효과적인 운동 강도를 규명하고자 하였다. 연구결과, 생쥐의 신체적 특성에서 그룹 간 유의한 차이는 나타나지 않았다. 생쥐의 운동 전과 후에 따른 젖산 변화에서도 유의한 차이가 나타나지 않았으며, 이는 초기 젖산의 높은 농도가 원인인 것으로 보인다. 이에 따른 선행연구들을 살펴본 결과, Ferreira et al. [23]의 연구에서, 생쥐의 운동 전 젖산은 2 mmol/L 로 나타났으며, 또한 Billat et al. [24]의 연구에서도 2, 3 mmol/L 정도로 나타났다. 우리의 연구에서 생쥐의 초기 젖산 농도는 4-6 mmol/L로 선행연구들에 비해 다소 높게 나타났으며, 이는 생쥐의 환경적 스트레스로 인한 젖산의 증가로 보인다. 본 연구에서는 선행연구와는 달리 생쥐의 트레드밀 운동 순응(acclimation) 기간을 두지 않았으므로, 이에 대해 생쥐의 초기 젖산 농도가 다소 높게 나타난 것으로 보이며 추후연구에서는 1주일 정도의 환경 적응기간을 두는 것이 타당한 것으로 사료된다.
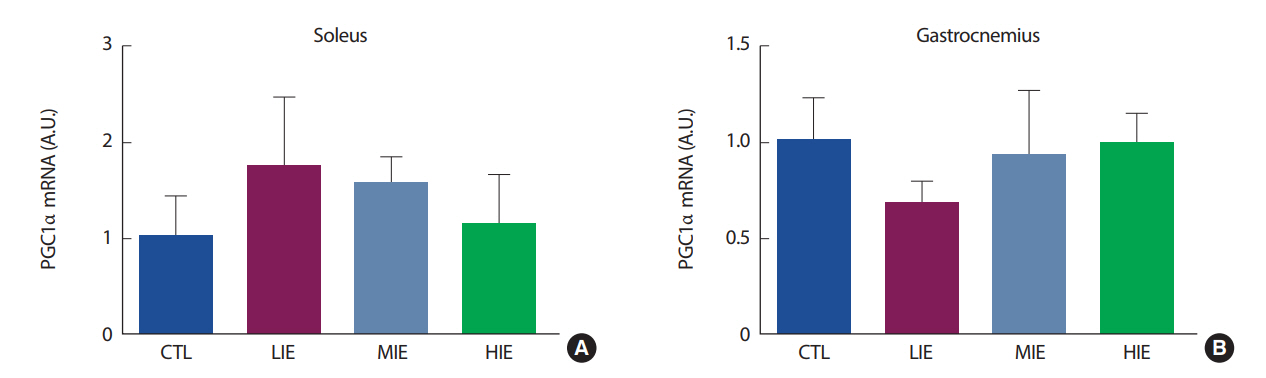
이 연구에서는 골격근에서의 PGC1α, FNDC5를 살펴보기 위해 전형적인 type I 섬유 유형을 가진 가자미근과 type I, type II 섬유의 특징을 모두 가진 비복근을 사용하였다. 연구 결과, 유산소 운동 강도에 의한 골격근에서의 PGC1α, FNDC5 변화에는 비복근에서 유의한 차이가 나타나지 않았다. 하지만, 가자미근에서 FNDC5 mRNA는 유산소 운동에 의해 유의한 변화가 관찰되었으며, 이는 근 섬유 유형에 따라 FNDC5 mRNA의 발현 정도에 차이가 있음을 나타낸다. FNDC5는 irisin이 혈액으로 방출되기 전 단계로[1], 유산소 운동 강도가 높을수록 증가하였으며 이는 우리의 가설과 일치한 결과를 나타냈다. Boström et al. [1]의 연구에 의하면, FNDC5는 PGC1α에 의해 조절되는 것으로 알려져 있다. 하지만, 우리의 연구에서는 가자미근에서 FNDC5 mRNA의 발현만 증가했을 뿐, PGC1α의 발현이 유의하게 증가하지 않아 선행연구와 다른 결과를 보여주었다. Pekkala et al. [14]의 연구에서도, 운동 후 FNDC5의 증가가 PGC1α의 증가를 유도하지 않는 것으로 나타났다. 이는 FNDC5의 증가가 반드시 PGC1α에서만 조절되는 것이 아닐 수도 있다는 것을 의미하며 따라서 FNDC5 발현에 관여하는 추가적인 기전에 대한 연구가 필요한 것으로 보인다.
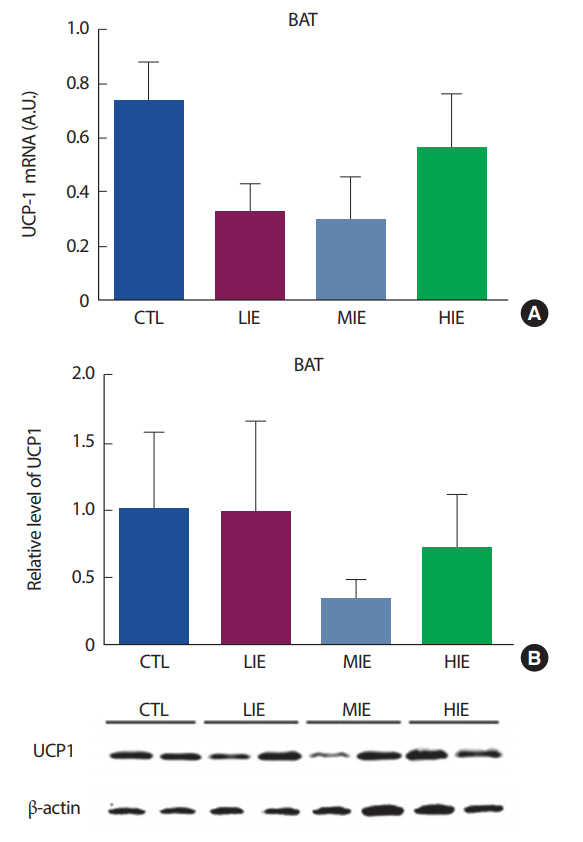
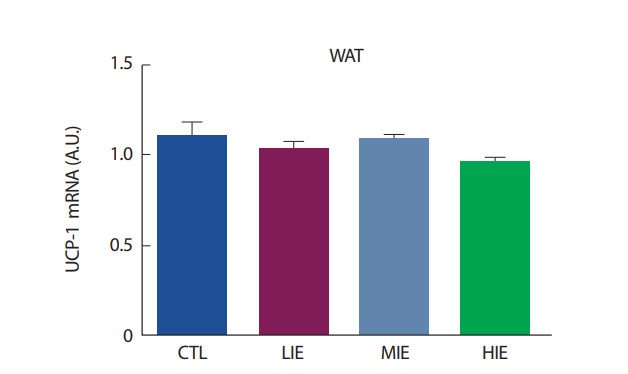
백색지방 조직은 크게 내장지방(visceral fat)과 피하지방(subcutaneous fat)으로 분류된다. 설치류 모델에서, 운동 훈련은 두 지방 조직에서 UCP1의 발현을 증가시켜 미토콘드리아 생합성을 증가시킨다. 하지만, 이러한 효과는 피하지방에서 훨씬 더 두드러진다[25]. 특히, irisin은 피하지방 조직을 자극하여 UCP1 발현을 증가시키는 것으로 알려져 있다[1]. 하지만, 우리의 연구에서는 백색지방조직을 생쥐의 부고환 주변 지방(epididymal fat), 즉 내장지방을 사용하여 분석이 이루어졌다. Ringholm et al. [26]의 연구에서, 1시간의 유산소 운동 후 생쥐의 내장지방과 피하지방을 추출하여 UCP1의 발현을 살펴본 결과, 운동 직후 피하지방에서의 UCP1이 내장지방의 UCP1보다 더 증가하는 것으로 나타났다. 또한, 운동에 의한 UCP1의 발현은 PGC1α의 발현이 증가해야 UCP1 또한 증가가 되는 것으로 나타났으며[26], 우리의 연구에서는 PGC1α의 발현 또한 유의하게 증가하지 않았으므로, UCP1이 유의하게 증가하지 않은 데에 영향을 미쳤을 것으로 보인다.
이 연구에서는 30분의 1회성 운동 강도별 유산소 운동이 운동 직후 irisin 증가에 유의한 영향을 미치지 않았다. 본 연구에서, 운동에 의한 FNDC5 mRNA의 증가는 가자미근에서만 나타났으며 이는 운동에 의해 Type I 근섬유에서 FNDC5의 자극이 이루어졌음을 나타낸다. 하지만, FNDC5 mRNA의 증가만 나타났을 뿐, FNDC5의 단백질 발현에서는 유의한 차이가 나타나지 않았다. 이는 30분의 일회성 유산소 운동이 FNDC5가 mRNA에서 단백질로 전달되기까지 충분한 자극이 이루어지지 않았으며, 이는 혈청 irisin이 유의하게 증가하지 않은 원인으로 보인다. Irisin은 FNDC5가 분해되어 혈액으로 방출되는 것으로 알려져 있다[1]. 우리의 연구에서, FNDC5의 단백질 발현에서는 유의한 차이가 나타나지 않았으므로, irisin이 혈액으로 방출되기까지 충분한 자극이 이루어지지 않은 것으로 사료된다.
이 연구에서는 운동 강도에 따른 그룹별 차이가 나타나지 않았다. 본 연구에서, 생쥐를 집단별 5마리씩 배정하여 비교적 적은 개체수로 연구를 실시하였고 전체적인 생쥐의 운동량에 대한 통제는 하지 못했다. 따라서 추후 연구에서는 운동량에 대한 정확한 통제와 생쥐의 개체수를 증가시키고 피하지방의 UCP1을 살펴봄으로써 운동 강도에 따른 irisin 발현의 차이와 이와 관련된 기전을 탐색하기 위한 추가 연구가 필요할 것으로 판단된다.
결 론
이 연구는 생후 13-15주 사이의 건강한 수컷 wild type C57BLKS/J-m+/m+ 생쥐를 대상으로 유산소 운동 강도에 따른 irisin 및 FNDC5 발현 정도의 차이를 밝히고자 하였다. 그 결과 1회성 유산소 운동으로 생쥐의 FNDC5 단백질과 irisin 발현에는 유의한 차이가 나타나지 않았으며, 이는 30분의 1회성 유산소 운동이 FNDC5 mRNA에서 단백질로 전달되기까지 충분한 자극이 이루어지지 않아 혈중 irisin 발현에도 영향을 미친 것으로 판단된다. 따라서 이 연구에서는 1회성 유산소 운동이 운동 강도에 따라 생쥐의 FNDC5 mRNA를 증가시키는 경향을 살펴봄으로써 FNDC5 mRNA가 유산소 운동 강도에 따라 증가할 것이라는 시사점을 제시하였다. 추후 연구에서는 인간을 대상으로 1회성 운동 강도에 따른 irisin 및 FNDC5 발현의 변화를 살펴봄으로써 대사 질환의 예방 및 완화에 운동이 어떻게 irisin 관련 기전을 통해 기여할 수 있는지를 이해할 수 있을 것이다.