


INTRODUCTION
Hypoxic training has gained widespread popularity among elite athletes as a means of enhancing endurance capacity. Hypoxic adaptation encompasses a series of responses aimed at acclimating to a low-oxygen environment, ensuring an adequate supply of oxygen molecules to the tissues. Key hypoxic responses include increased ventilation, cardiac out-put, erythropoiesis, glycolysis, and angiogenesis [1]. Athletes exposed to hypoxic adaptation responses often witness improvements in endurance exercise performance, attributed to increases in red blood cells, hemoglobin, capillaries, and a shift in muscle fiber type towards the slower twitch.
Hypoxia-inducible factor (HIF) emerges as a critical transcription factor responsive to low oxygen tension at the cellular level [2-4]. The HIF comprising α and β subunits are hydroxylated by prolyl hydroxylase domain proteins (PHDs), allowing ubiquitination by the VHL H3 ubiquitin ligase leading to rapid degradation by the proteasome under normoxic conditions [5,6]. In hypoxic conditions, HIFs stabilize and trans-locate into nuclear by inhibiting prolyl hydroxylase. HIF binds to Hy-poxia-responsive elements (HRE) to activate target genes related to erythropoiesis, glycolysis, and angiogenesis [7,8]. Among the PHDs iso-form (PHD1, 2, and 3), PHD2 serves as the primary regulator of HIF-1α hydroxylase, while PHD1 and 3 mainly play a compensatory role under hypoxic condition [5,9-12].
Indeed, studies shown that HIF-1α modulated hypoxic ventilatory responses during hypoxic training in athletic players [13]. Furthermore, investigations with HIF-1α overexpressed mouse model, induced by the PHD2 deficiency demonstrated an increase in slow fiber-type twitch and capillary density in skeletal muscle. PHD2KO mice exhibited a more pronounced effect from an 8-week endurance training regimen compared with WT littermate mice [14,15]. Additionally, HIF-1α regu-lates glycolytic metabolism to produce ATP in conditions of insufficient oxygen concentration, making glycolysis crucial during high-speed or resistance exercise performance [16,17]. However, it remains unclear whether the upregulation of glycolysis induced by the HIF-1α dependent pathway improves exercise capacity. Overall, this study aims to elucidate the effects of the hypoxic-responsive pathway on exercise performance using the PHD2-deficient mouse model. This mouse model enables to stabilization of HIF-1α by genetically inhibited PHD2 under normal oxygen level conditions. Therefore, the aim of this study is to verify that the hypoxic-responsive pathway, achieved through PHD2 inactivation, enhances exercise performance regardless of oxygen levels.
METHODS
1. Ethical approval
All animal experimental procedures were performed according to the protocols approved by the Guidelines for the Care of Laboratory Animals of Tohoku University Graduate School of Medicine (Sendai, Japan).
2. Animals
All experiments were conducted following the regulations of the Stan-dards for Human Care and Use of Laboratory Animals of Tohoku University (Permit Number: 2015ikokumikae-007, 2015ikodo-008). Because constitutive knock-out of Phd2 is embryonically lethal (Takeda et al. 2006), we used an estrogen receptor (ER) agonist-induced Phd2 knock-out mouse using the Cre-loxP system. Phd2/flox mice (a gift from Dr. G.H. Fong) were mated with Rosa26/Cre-ER (T2) mice to generate Phd2 conditional knock-out (KO) mice (Phd2f/f/Rosa26CreERT2; Phd2KO) by referring to a previous report (Takeda et al. 2007). To delete the floxed Phd2 exon 2, the ER agonist tamoxifen (Sigma, St. Louis, MO, USA) was delivered to 8- to 12-week-old male Phd2 KO mice by intraperitoneal in-jection (10 mg/mL in corn oil, 20 mg/kg/day for five consecutive days). Littermate male Phd2f/f mice lacking Cre were used as controls and were similarly treated with tamoxifen. For genotyping, DNA was isolated from tails. The skeletal muscle was harvested after treadmill exercise capacity test.
3. Maximal exhaustion exercise test
A running test using a rodent treadmill was performed 2 weeks after tamoxifen treatment to determine the exercise capacity. Before the test, the mice ran for 5 min at 10 m/min as a warm-up. The speed was set at 10 m/min and was increased by 2 m/min every min. Throughout the warm-up and test running, the treadmill was set fixed slope of 5°. The mice were encouraged to run by tail stimulation with a soft brush and a low-voltage power grid. Exhaustion was determined to be the point at which the animal would not resume running for 15 seconds despite gentle brushing on the tail and a mild electrical foot shock [18,19].
4. Blood profile
Whole blood was collected 6 weeks after tamoxifen administration from the control and Phd2 cKO mice via the buccal vein under anesthesia. All blood components were measured using a multiple automatic blood cell counter for animals (MICRO abc LC-152, Horiba, Tokyo, Japan).
5. Measurement of lactate concentration
Blood lactate concentration was measured by Arkray Lactate Pro 2 both right before and after treadmill exercise capacity test, using the mouse tail.
6. Immunoblot
To isolate total protein extracts, 50 mg of skeletal muscle tissue was homogenized for 30 s on ice in 1 mL of lysate buffer (40 mM Tris (pH 7.5), 300 mM KCl, 1% Triton X-100, 0.5 M EDTA, Protease inhibitor cocktail X 20 (Sigma)), using a Polytron PT-MR 2100 homogenizer. Ho-mogenates were centrifuged at 12,000 rpm for 5minutes at 4°C, and the supernatants were isolated. Protein concentrations were determined using the BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL) with bovine serum albumin (BSA) as the standard, and extracts were stored at −80°C. Nuclear and cytoplasmic extractions were performed using the NE-PER nuclear and cytoplasm extraction reagents (78833, Thermo Fisher Scientific). Total protein was separated via 8-12% SDS-PAGE and transferred to a PVDF membrane (Invitrogen). The membrane was blocked using Tris-buffered saline with 0.05% Tween 20 (TBST) containing 5% BSA for 1 hour and incubated overnight with ap-propriately diluted (1:500-1,000) primary antibody in TBST at 4°C. After incubation, the membranes were rinsed three times in TBST for 5 min-utes and incubated with secondary antibody in 4% skim milk for 1 hour at room temperature. Protein bands were visualized and quantified using a Molecular Imager VersaDoc 5000MP system (Bio-Rad) and ECL.
7. Cell culture
C2C12 myoblasts were cultured with high-glucose Dulbecco's modi-fied Eagle's medium (DMEM) supplemented with 10% fetal bovine serum and 100 µg/mL penicillin-streptomycin. The cells were transfected with 50 nM of PHD2 and scramble as negative control small interfering RNA (siRNA) using Lipofectamine RNAimax (Invitrogen) for 24 hours. Then, the siRNA was removed and washed with PBS. The transfected C2C12 myoblast was harvested at 24 hours after transfection.
8. RNA sequencing analysis
PHD2 siRNA transfected C2C12 myoblasts were lysed by RNA lysis binding buffer, and then stored at −80°C until RNA was extracted. RNA was isolated using the RNAeasy kit (Qiagen) following the manufacturer's instructions. RNA quality and quantity were determined using nanodrop (ThermoFisher). Total RNA (2 µg) was used for poly(A) selection to construct the RNAseq library. The libraries were sequenced on the Illumina HiSeq (2×150 bp). Sequence reads were trimmed to remove possible adapter sequences and nucleotides with poor quality using Trimmomatic v.0.36. The trimmed reads were mapped to the Mus mus-culus GRCm38 reference genome available on ENSEMBL using the STAR aligner v.2.5.2b. The STAR aligner is a spliced aligner that detects splice junctions and incorporates them to help align the entire read sequences. The splice variant hit counts were extracted from the RNAseq reads mapped to the genome to estimate the expression levels of alternatively spliced transcripts. Differentially spliced genes were identified for groups with more than one sample by testing for significant differences in read counts on exons (and junctions) of the genes using DEXSeq. The exon hit count tables were provided for groups with only one sample. The results of the splice variant expression analysis are included in sepa-rate reports.
RESULTS
1. PHD2KO mice result in a decrease in body weight and muscle wasting
To inhibit the activation of PHD2 in the adult stage, we employed PHD2 f/f Rosa26 CreERT2 mice. The tamoxifen was administrated at 8-12 weeks of age, and we validated the expression of PHD2 in gastrocnemius muscle post-tamoxifen administration through immunoblot analysis. We confirmed that the PHD2 expression was dramatically decreased and HIF-1α expression was increased in the gastrocnemius muscle of PHD2KO mice compared with PHD2 f/f littermate mice (Fig. 1A). Also, Additionally, PHD2KO mice showed a significant decrease in body weight and gastrocnemius muscle weight compared to PHD2 f/f littermate mice (Fig. 1B and C). Furthermore, blood profile analysis from fa-cial vein samples during anesthesia revealed that PHD2KO mice exhibited significantly higher red blood cell volume (RBC), hemoglobin, and hematocrit levels compared to PHD2 f/f mice (Table 1).
Fig. 1.
Fig. 1.The loss of Prolyl hydroxylase domain 2 results in muscle wasting. (A) Representative western blotting shows protein levels of PHD2 and HIF-1a from gastrocnemius muscle of PHD2f/f littermate wild-type and PHD2KO mice. (B) Body weight (C) Gastrocnemius muscle weight normalized by body weight. (n=8-9) Data are shown as mean SEM. **** p<.0001.

2. PHD2 deficiency promotes maximal exercise capacity and lower blood lactate concentration
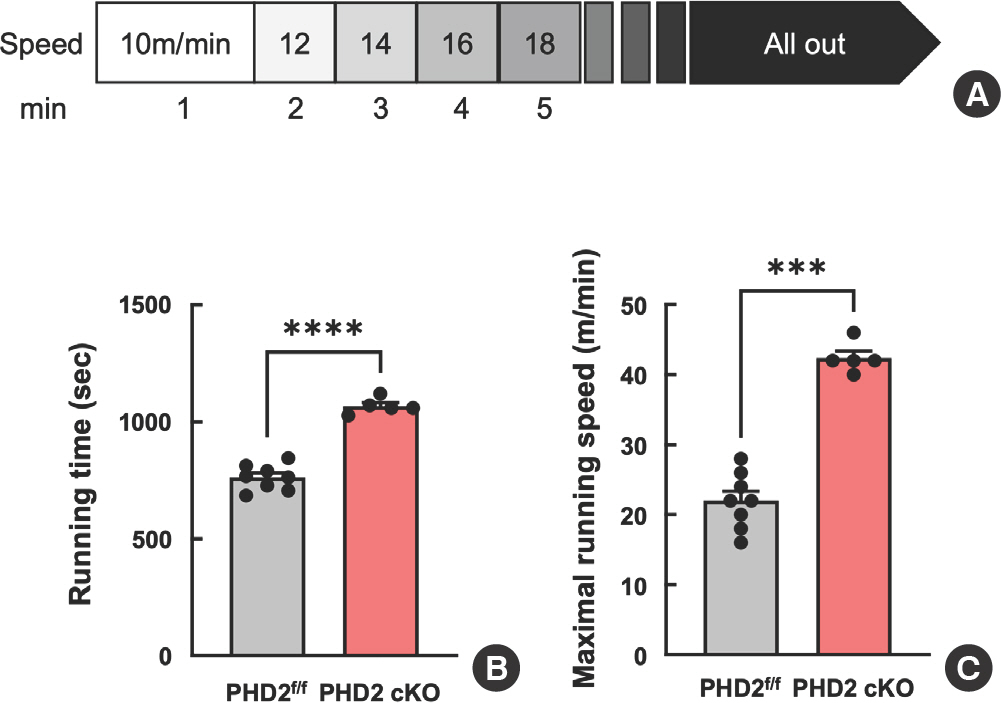
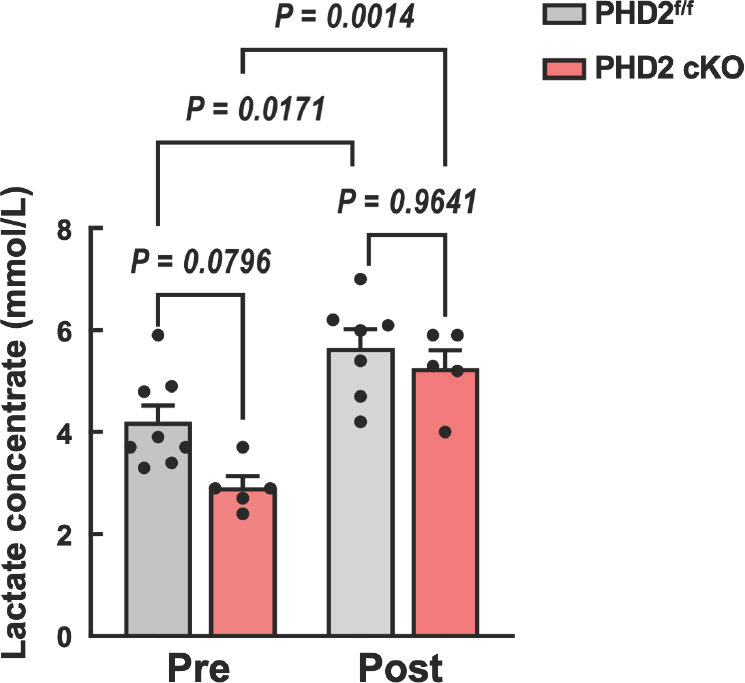
To investigate the impact of the hypoxic response pathway on exercise performance, we conducted a speed running treadmill test using PHD2 f/f and PHD2KO mice. The running speed was initiated at 10 m/min, in-crementing by 2 m/min every minute with a fixed slope of 5° (Fig. 2A). Notably, PHD2KO mice had higher running time and maximal running speed compared with PHD f/f littermate WT mice (Fig. 2B, C). Furthermore, blood lactate level was measured at pre- and post-running tests. PHD2KO mice showed a lower lactate level at the pretest com-pared with PHD2 f/f littermate WT mice. However, no significant difference in lactate levels was observed between PHD2KO and PHD2 f/f mice after running the test (Fig. 3A).
Fig. 2.
Fig. 2.Prolyl hydroxylase domain 2 deficiency enhanced maximal exercise capacity. (A) The schematic design of an experimental protocol for the maximal exercise capacity test. The speed was set at 10 m/min and was increased by 2 m/min every minute with a fixed slope of 5o. (B) Average running time (sec) and maximal running speed in treadmill exercise capacity test (n=5-8). Data are shown as mean SEM. *** p<.0005, **** p<.0001
3. RNA sequencing demonstrates that PHD2-deficient muscle cell increases the expression of genes related to glycolysis and muscle contraction
To investigate the role of the PHD2/HIF-α-dependent pathway in muscle glycolytic metabolism, we utilized a PHD2 knockdown model through siRNA transfection in C2C12 myoblasts to exclude the effect from other cells. Because the skeletal muscle tissues consist of various cell types including vessels (Smooth muscle cells, endothelial cells, and pericytes), connective tissues, and myocyte. Immunoblot analysis confirmed successful PHD2 knockdown and increased HIF-1α expression in siPHD2 compared to scramble-transfected myoblasts (Fig. 4A). Interestingly, RNAseq analysis demonstrated a significant upregulation of key glycolysis-related genes in siPHD2 myoblasts compared to the control. Specifically, 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 enzyme (PFKFB3), Phosphoglycerate mutase 1, and 2 (PGAM1 and 2), 5’-AMP-activated protein kinase catalytic subunit alpha-2 (Prkaa2), and Hexokinase-1, and 2 (HK1 and 2) were dramatically increased in PHD2-deficient myoblast compared with control. Furthermore, genes associated with muscle contraction including calsequestrin (Casq1), troponin 1 and 3 (Tnni 1 and 3), and alpha-actinin-3 (Actn3) were significantly up-regulated in PHD2-deficient myoblast compared to the control. (Fig.4B).
Fig. 4.
Fig. 4.Loss of PHD2 in skeletal muscle increase glycolysis and muscle contraction-associated gene expression. (A) Representative immunoblot for PHD2 and HIF-1α in C2C12 myoblast with PHD2 siRNA transfection. (B) Heat map of RNAseq analysis showed significantly up-regulated (red) or down-regulated (blue) genes related to glycolysis and muscle contraction from scramble and siPHD2 transfected C2C12 myoblast. Scramble: n=3; PHD2KD: n=3.

DISCUSSION
In this study, we demonstrated that the activating glycolytic metabolism via the HIF-α pathway enhances exercise capacity utilizing PHD-2KO mice. The main findings of the present study were that the hypox-ic-responsive pathway by PHD2 deficiency 1) increased red blood cell (RBC) and hemoglobin levels, 2) had lower blood lactate levels, 3) acti-vated the gene expression related to glycolytic metabolism and muscle contraction, 4) enhanced exercise capacity under normoxic and normo-baric condition. This finding is the first evidence that the HIF-α path-way improves exercise capacity without exercise training.
Traditionally, hypoxic training has primarily focused on improving endurance exercise performance in elite athletes. Our previous studies demonstrated that the elevation of RBC and hemoglobin levels, coupled with muscle fiber type transition towards slow twitch and increased muscle capillary density through the HIF-α responsive pathway via PHD2 deficiency, led to enhanced effects of endurance exercise training compared with WT littermate mice [14,15]. The RBC and hemoglobin are elevated by the upregulation of the Epo gene in Epo-producing cells (REPC) located in the cortex and outer medulla of the kidney under the HIF-α pathway [20]. Also, we observed that PHD2KO mice decreased body and muscle weight compared with WT littermate mice. The muscle wasting in PHD2KO mice might be originated from the decrease in fast fiber twitch and an increase in the gene expression related to muscle atrophy such as Murf1 and atrogin-1 [15]. Moreover, hypoxic exposure has been associated with a reduction in myotube diameter and increased muscle atrophy markers, highlighting the complexity of the interplay between hypoxia and muscle metabolism [21].
This study revealed that PHD2 deficiency results in a reduced blood lactate concentration during steady-state conditions compared with wild-type (WT) mice. Lactate, an end product of glycolysis, is a waste product associated with muscular fatigue during exercise. Recent evidence indicates that lactate produced from pyruvate in glycolysis can be converted to glucose via the Cori cycle in the liver. Minamishima demonstrated that inhibiting PHD2 activates the lactate-glucose carbon re-cycle system through the Cori cycle, leading to a reduction in blood lactate concentration [22]. Notably, liver-specific-PHD2KO mice showed lower blood lactate levels after 50-minute treadmill exercise compared to control mice [22]. However, our mice did not show a significant decrease in lactate concentration after the maximal running test. This outcome suggests potential benefits for exercise performance and warrants fur-ther investigation into the nuanced effects of PHD2 deficiency on lactate metabolism during different types of physical activity.
To gain better insight into muscle metabolism under the hypoxic-re-sponse pathway, we performed RNAseq analysis using C2C12 myoblast which is widely used for mouse skeletal muscle cell for in vitro experiments. The inhibition of PHD2 by siRNA transfection resulted in a significant upregulation of genes related to glycolysis and muscle contraction. Glycolysis, a fundamental process in rapidly providing ATP to contracting skeletal muscles during anaerobic exercise, plays a crucial role. The glycolytic pathway's ability to produce ATP at a faster rate than oxidative phosphorylation is essential for meeting the energy demands of intense physical activity [17,23,24]. Moreover, our analysis highlighted the increased expression of key regulators involved in calcium handling during muscle contraction. Calsequestrin (Csaq1), the primary Ca2+ binding protein within the sarcoplasmic reticulum, along with Sarco-plasmic/Endoplasmic Reticulum Calcium ATPase 1 and 2 (SERCA1, 2), emerged as crucial players in Ca2+ storage and handling in contracting skeletal muscles [25-27]. Notably, the heightened expression of Csaq1 and SERCA1 was observed after sprint running training in the gastrocnemius muscle. The increased expression of genes related to muscle contraction, such as Csaq1, SERCA1, Tnni1, and Actn3, is widely recognized as beneficial for anaerobic exercise performance. These findings from the RNAseq analysis underscore the pivotal role of metabolic changes induced by the hypoxic-responsive pathway in actively contracting muscles, providing valuable insights into the molecular mechanisms sup-porting enhanced exercise performance.
CONCLUSION
The present study provides novel evidence demonstrating that the hypoxic response pathway by PHD2 inhibition enhances the gene expression related to glycolysis, gluconeogenesis, and exercise capacity. These findings imply that the metabolic changes induced by the PHD2/HIF pathway may contribute to improved exercise performance. Furthermore, the metabolic shift in skeletal muscle could pave the way for the future translational studies aimed at preventing metabolic disorders such as diabetes or atherosclerosis.


