서 론
지방조직으로의 과도한 지질 축적은 지방 대사에 문제를 일으켜 심각한 대사 장애를 유발하는 “에너지 항상성(energy homeostasis)”의 파괴를 일으키는 것으로 널리 알려져 있다[1-3]. 지방조직은 지질의 저장소로서 역할을 하며 칼로리 섭취량과 소비량이 적절히 유지되면 대사장애가 발생할 수 있는 가능성이 희박하다. 그러나 에너지 소비량이 감소하고 반대로 섭취량이 증가하게 되어 균형이 깨질 경우 지방세포에서 에너지 저장이 증가하고 비대해진다[1,2,4,5]. 비대해진 지방세포는 nucleotide-binding oligomerization domain-like receptors (NOD-like receptors)의 아족(subfamily)으로 염증 경로의 수를 활성화시키는 NOD-leucine rich repeat and pyrin domain containing 3 (NLRP3) 인플라마좀(inflmmasome)의 활성화와 같은 몇 가지 신호 결함을 유발하기도 한다[6]. NLRP3는 에너지 항상성 유지 경로 중 sirtuin 1 (SIRT1)로를 손상시킴으로써 인슐린저항성과 제2형 당뇨를 일으키는 염증의 진행에 크게 기여한다고 알려져 있다[7-9]. 또 다른 에너지 센서는 AMP-activated protein kinase (AMPK)가 있다[7,10]. AMPK는 비만으로 비롯된 과량의 지질이 아디포텍틴 수치를 감소시켜 아디포넥틴에 의해 조절되는 AMPK가 불활성화되어 SIRT1과 함께 에너지 대사 결함을 일으키는 주요 원인이 된다[11,12].
에너지 항상성은 세포의 에너지 센서인 SIRT1과 AMPK로 인해 유지되며 세포 에너지의 고갈을 감지하는 역할을 한다. 앞서 언급한 바와 같이 비만으로 인한 NLRP3의 활성화는 SIRT1을 비활성화시켜 에너지 센싱을 무디게 한다. 따라서 에너지 고갈이 감지되지 못하여 축적된 에너지 사용을 막고 지방을 축적시킨다. SIRT1은 NAD+의존적인 탈아세틸화 효소이며, NAD+의 증가는 SIRT1을 활성화시키는 반면, AMPK는 AMP/ATP 비율이 증가할 때 활성화된다[7,8,11,12]. 이 단계에서 SIRT1은 PGC1α를 탈아세틸화하는 반면 AMPK는 PGC1α를 인산화시키고 과다한 유전자를 발현하는 불활성 PGC1α를 활성화시키며, 그중 대부분은 에너지 생산에 있어 지질의 동원에 관여한다[7,13,14]. 이러한 모든 과정은 주로 지방세포에서 발생한다. SIRT1과 AMPK 경로는 에너지 항상성 유지에 매우 중추적인 역할을 하며 해당 경로의 비활성화는 대사적 결함을 일으킬 수 있는 주요 경로가 될 수 있다.
Fetuin A는 헤파토카인(hepatokine)으로 잘 알려져 있었다. Fetuin A는 대사성 질환에서 주요 병리학적 역할을 하는 것으로 입증된 최초의 헤파토카인으로 당단백(glycoprotein)이다[15]. Fetuin A는 지방증(steatosis)과 염증이 유발된 간(liver)에서 증가하는 것으로 알려졌다[15]. 주로 간에서 분비되어 혈중 fetuin A의 수치가 높으면 제2형 당뇨병이 발생할 위험이 높고[16] 심근경색[17]과 뇌경색의 발생위험이 높다고 보고되어 있다[18]. 그러나 최근 새롭게 alpha-2-HS-glycoprotein (fetuin A)이 지방조직에서 합성되는 것이 알려졌으며[19], 에너지 센서인 SIRT1과 AMPK 경로를 모두 손상시켜 에너지 항상성의 파괴를 초래한다는 것이 밝혀졌다[20]. 따라서 우리는 운동이 fetuin A의 억제에 얼마나 기여할 수 있는지 의문을 가지게 되었다.
문헌조사 결과 운동과 관련하여 혈장 순환 수준에서 관찰된 연구 중 사람을 대상으로 운동중재를 실시한 연구가 총 7건 있었다[21-27]. 이 중 4건의 연구에서 순환 수준에서의 혈장 fetuin A가 감소한 결과로 나타났다[21,22,24,26]. 그러나 나머지 3건의 연구에서는 아무런 차이가 나타나지 않았다. 사람에서의 이러한 운동에 의한 효과가 일치하지 않는 것은 단지 혈장 fetuin A뿐만 아니라 지방조직에서의 발현 또한 중요한 의미를 지닐 수 있음을 시사하기도 하며 혈장에서와 지방조직에서의 반응이 충분히 다를 수 있고 그것이 중요한 의미를 가질 수 있음을 예측 가능하게 한다. 최근의 논문[28]에서 언급하고 있듯이 지방조직이나 간이 순환 fetuin A에 상대적으로 기여할 수 있다는 것을 밝혀야 함에도 불구하고 현재까지는 운동 중재 후에 간이나 지방조직에서 변화하는 fetuin A를 관찰한 연구는 극히 드물다. 이러한 이유는 계획되고 설계된 통제 내에서 중재를 하고 그에 대한 분자생물학적 실험을 위해 지방조직이나 간을 생검(biopsy)하는 것은 연구윤리위원회에 의해 기각을 받을 확률이 매우 높고 또한 침습적인 실험방법으로 인해 연구에 충분한 대상자를 확보하기가 쉽지 않기 때문인 것으로 생각된다. 이처럼 fetuin A 관련 연구들은 운동관련 연구를 포함하여 사람을 대상으로 혈장 수준에서의 평가가 이루어진 연구가 대부분이었다. 그러나 fetuin A가 지방조직에서 SIRT1과 AMPK 경로를 억제한다는 연구[20]가 2016년에 보고되었기 때문에 관련 연구가 부족한 것은 당연한 것으로 보여진다.
우리는 운동이 지방조직의 fetuin A에 영향을 미칠 수 있다는 가능성을 Perez-Soleto와 그의 동료들의 연구[29]를 통해 확인하였다. 해당 연구에서 랫드(rat)와 세포 주 실험을 병행하여 다양한 대사적 상황에서의 fetuin A의 분비를 평가하였다. 그러나 연구의 목적이 내장과 피하지방의 상황에 따른 fetuin A 분비 차이를 확인하고자 함이였고 실제로 연구에서 운동은 실제 비만하지 않은 쥐에서 수행되었다. 해당 연구의 결과에서 비만에 의해 내장의 백색 지방 조직(white adipose tissue)에서 fetuin A가 유의하게 증가한다는 것이 확인되었지만, fetuin A가 금식과 자발적 운동 훈련 후에 감소한다는 것은 정상 쥐를 대상으로 확인되었고 비만 쥐를 대상으로 fetuin A가 운동으로 인해 감소될 수 있는지는 확인되지 않았다. 결론적으로 fetuin A가 운동을 통해 비만인 사람이나 비만 동물의 지방조직에서 억제할 수 있는지는 확인된 바가 없다. SIRT1와 AMPK에 의한 에너지 항상성은 주로 지방조직에서 이루어지고 비만에 의해 fetuin A가 실제로 증가하는 것으로 알려져 있기 때문에 이를 운동을 통해 감소시킬 수 있는지는 확인이 필요하다. 이를 위해서는 지방조직에서 지방세포를 추출하여 fetuin A를 분석하여야 한다.
우리는 세포의 에너지 항상성에 관여하는 SIRT1과 AMPK 모두 급성 운동에 의해 신호 전달을 증가[30]시킬 수 있는 것과 마찬가지로 비만에 의해 증가된 fetuin A를 일회성의 운동으로도 억제하고 인슐린 감수성을 증가시킬 수 있을 것이라고 생각하였다. 실제로 일회성 운동에 의해서 fetuin A가 감소한다면 운동으로 인해 지방조직에서의 에너지항상성이 정상적으로 조절할 수 있다는 것을 의미므로 지방조직에서의 염증반응 및 에너지 센싱 감도를 높이는 직접적인 작용을 한다는 것이 확인되는 것이다. 따라서 장기간의 규칙적인 운동은 fetuin A를 억제함으로써 SIRT1 또는 AMPK 경로의 활성화를 유도하여 지방조직 내의 에너지 항상성을 유지하는 데 기여를 할 수 있다는 기대를 갖게 하고 또한 사람에게 장기간의 규칙적인 운동을 제안하는 중요한 근거가 될 것이다.
본 연구의 목적은 일회성의 운동만으로도 fetuin A가 지방조직에서 억제될 수 있다는 사실을 새롭게 밝히는 것이며, 또한 이전의 fetuin A 관련 연구들이 대부분 혈장에서 연구되어 있기 때문에 fetuin A의 지방조직에서의 분비와 혈장 순환 수준에서의 농도와의 관련성을 확인하고자 하였다.
이를 위해 우리는 고지방식이로 유도된 비만쥐를 대상으로 일회성의 운동 후 지방조직에서 지방세포를 추출하여 fetuin A의 단백질 발현을 측정하였고 혈장에서의 fetuin A 농도를 측정하여 상관관계를 분석하였다.
연구 방법
1. 연구 설계
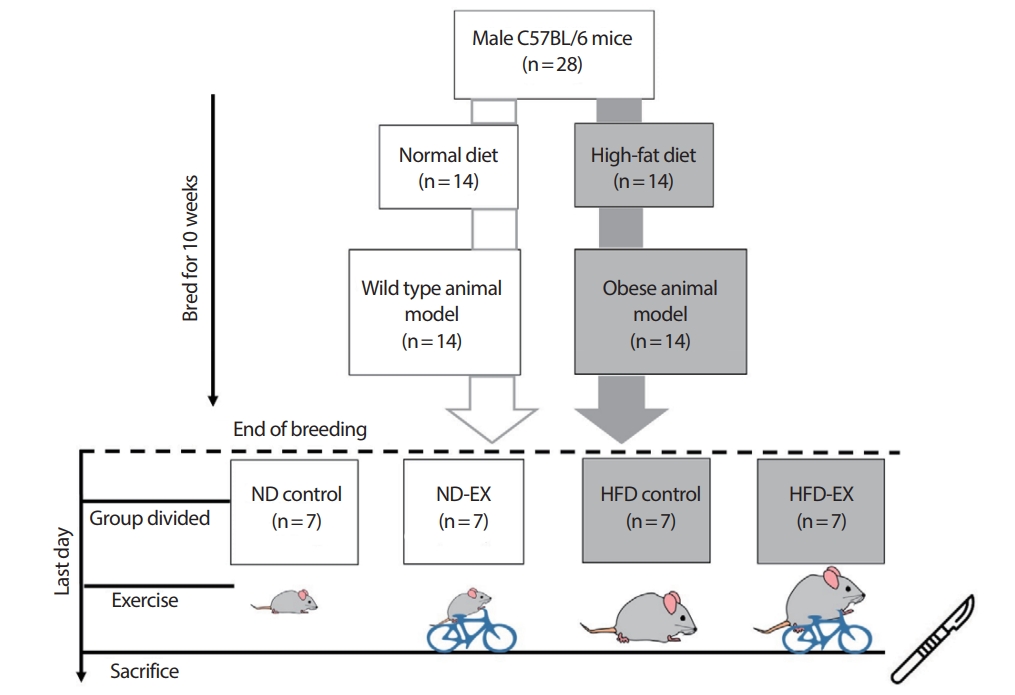
6주령의 수컷 C57BL/6 종 마우스 28마리를 분양받아 10주간 사육되었다. 실험기간 동안 실험동물 중 14마리는 일반식이를 섭취하였으며 나머지 14마리는 고지방식이를 섭취하여 비만이 유도되었다. 10주간의 사육이 끝난 후 일반식이를 섭취한 실험동물 중 7마리와 고지방식이를 섭취한 실험동물 중 7마리를 각각 무작위 선별하여 실험 종료일에 일회성 운동을 실시하였다. 따라서 최종 집단 구성은 일반식이군(normal diet control, ND control; n=7), 운동군(ND exercise, ND-EX; n=7), 고지방식이군(high-fat diet control, HFD control; n=7) 그리고 고지방운동군(HFD exercise, HFD-EX; n=7)으로 이루어졌다(Fig. 1). 실험동물은 자동 항온 및 항습 그리고 12시간 자동 명암주기 시스템이 설치된 동물실험실에서 사육되었다. 또한 식이와 음수는 자유식이(ad libitum)로 공급되었다. 본 연구는 한 번의 동물실험으로 파생된 실험의 일부분으로 본 운동과학 학회지에 게재된 연구인 “고지방식이로 유도된 비만 마우스의 지방조직에서 일회성 운동이 대식세포 표현형 비율 변화에 미치는 영향”과 동일한 실험설계와 동일한 조직의 일부를 사용하였음을 밝히며 공통적으로 사용된 데이터에 대해서는 이에 대한 사실을 알림과 함께 출처를 표시하였다[31]. 본 연구는 부산대학교 동물실험윤리위원회의 승인(PNU-2017-1669)을 받아 실시되었다.
2. 운동 방법 및 체중 및 식이섭취량 측정
모든 일회성 운동 집단은 동물용 트레드밀(DJ-344; Daejong Instrument Industry, Daejeon, Korea)에서 운동을 수행하였다. 마우스의 운동강도는 C57BL/6 마우스의 임계속도인 18.7 m/min을 초과하지 않도록 운동강도를 설정하여 운동 이외의 스트레스를 최소화하였다[32]. 운동강도 설정을 위해 최종적으로 일회성 운동을 실시하기 2주 전에 하한 속도 12 m/min, 상한 속도 18.7 m/min 사이에서 예비 실험을 실시하였다. 예비 실험 결과 15 m/min의 속도가 정상 마우스와 비만 마우스 모두 트레드밀에서 뒤처지지 않고 60분간의 운동을 수행할 수 있는 최적의 운동 강도임을 확인하였다. 예비 실험 결과를 바탕으로 일회성의 운동은 트레드밀 작동 후 5분간 5 m/min의 속도로 warm-up을 실시하고 이후 60분 동안 15 m/min의 속도로 운동을 수행하였다. 운동 수행이 끝난 후 5분 동안 5 m/min의 cool-down을 실시하였다. 이때 운동을 실시하지 않는 실험동물들도 운동 시작과 동시에 식이공급을 중단하고 운동 실시 중에 운동 수행군과 같은 공간에 배치하여 트레드밀의 소음과 진동을 느끼도록 하였다. 식이섭취량과 체중은 실험기간 동안 주 1회 측정되었다.
3. 분석 방법
1) 조직 및 혈액 채취
연구목적에 따라 일회성 운동에 대한 운동과 식이의 영향을 모두 포함하기 위하여 10시간 이상의 절식은 실시하지 않았다. 장시간의 절식은 에너지 고갈과 관련하여 SIRT1과 AMPK의 발현 및 인슐린저항성에 관여하는 염증 관련 유전자의 발현에 영향을 미칠 수 있다[33]. 따라서 절식은 최소화하고 식이 섭취 후 혈당과 인슐린의 수치가 기저치로 돌아올 수 있도록 단시간의 절식을 실시하였다. 절식 동안 음수는 정상적으로 공급되었다. 실험동물은 이산화탄소 흡입을 통하여 희생되었으며 즉시 1 mL 주사기를 실험동물의 흉골 밑으로 삽입하여 심장의 혈액을 채취하였다. 이어서 복부를 절개하여 대표적인 WAT인 부고환 지방조직을 채취하였다. 채취한 혈액은 4 mL EDTA튜브(BD Diagnostics, Plymouth, UK)에 담아 13,000 rpm에서 원심분리하여 상등액인 혈장만 분리되었고, 1 mL 테스트 튜브에 옮겨 담았다. 또한 지방조직은 0.9% 생리식염수로 세척하여 12-well plate에 나누어 담았다. 혈액과 지방조직은 분석에 이용하기 전까지 -80°C 초저온 냉동고에 저장되었다.
2) 경구당부하 검사(oral glucose tolerance test, OGTT)
비만으로 인한 대사적 이상이 발생하였는지 확인하기 위하여 OGTT를 실시하였다. OGTT는 실험 종료 1주일 전에 실시하여 OGTT 과정에서 실시되는 절식에 대한 영향을 피하고자 하였다. 총 17시간의 절식 후에 OGTT가 실시되었다. 절식 시작 전 깔집이나 사료 부스러기가 없는 깨끗한 케이지로 실험동물을 옮긴 후 절식을 실시하여 공복 혈당의 개입요소들을 완벽히 차단하였다. 음수는 정상적으로 공급하였다. 꼬리정맥(tail vein)을 면도날로 약하게 베어내 혈액을 채취하고 공복 시 혈당을 측정하였다. 이후 20%의 포도당 수용액을 곡선 위관(curved gavage needle) 삽입을 통하여 경구투여하였으며, 투여량은 절식 후 측정된 공복 체중에 근거하여 산정되었다(공복시 체중[g]×10 =20% 포도당 수용액 투여량[μL]). 포도당 수용액의 경구투여 후 각 15분, 30분, 1시간 그리고 2시간 후에 맞추어 꼬리정맥을 가볍게 압박하여 맺힌 혈액을 측정지에 점적하여 측정되었다. 모든 혈액은 ACCU-CHEK® active (Roche Diabetes Care GmbH, Mannheim, Germany) 혈당측정기(blood glucose meter)를 이용하여 측정되었다. 포도당 제거율(glucose clearance)을 평가하기 위하여 시간 경과에 따른 혈당치를 측정하여 꺾은선 그래프로 나타내어 곡선하면적(area under the curve, AUC)을 삼각함수법(trigonometric function)으로 계산하였다[34].
3) Western blot
분리된 지방조직은 메스로 잘게 자른 다음 차갑게 준비된 RIPA 완충액(200 mM Tris/HCL [pH 7.4], 130 mM NaCl, 10% [v/v] glycerol, 0.1% [v/v] SDS, 1% [v/v] Triton X-100, 10 mM MgCl2)으로 융해시켰다. 융해된 지방조직은 4°C에서 13,000 rpm으로 10분간 원심분리하여 단백질이 추출되었다. 추출된 단백질은 Bradford’s assay로 정량분석된 동일한 양의 단백질(50 μg/well)을 sulfate-polyacrylamide gel (SDS-PAGE)에 분주하고 200 V에서 35분간 polyvinyl fluoride (PVDF) membrane에 transfer하였다. Transfer가 완료된 membrane은 5% 탈지분유(skim milk)로 1시간 동안 blocking을 실시하였다. 이후 1차 항체인 anti-AHSG antibody (1:2,000, Abcam, Cambridge, MA, USA)를 넣고 4°C에서 10시간(overnight) 동안 반응시켰다. 이후 2차 항체인 goat anti-rabbit IgG H&L (HRP) (1:100,000, Abcam, Cambridge, MA, USA)을 결합시켰다. 실험이 완료된 membrane은 enhanced chemiluminescence (ECL) 용액(Bionote, Hwaseong, Korea)으로 1분간 반응시킨 후 cassette에 넣고 투명필름으로 덮고 다시 X-ray 필름(Ortho CP-G Plus flim, Agfa-Gevaert NV, Belgium)을 포개어 5초간 감광시켰다. 형광에 노출된 X-ray 필름은 developer 용액과 fixer 용액으로 현상하여 밴드를 확인하였다. 밴드의 픽셀 강도를 FluorChem HD2 (Alpha Innotech, San Leandro, CA, USA) 프로그램으로 분석하여 발현량이 측정되었다. Fetuin A의 발현수준은 GAPDH 수준에 대비하여 결정되었고 대조군과 비교하여 표준화(normalization)되었다.
4. 자료처리방법
통계분석은 GraphPad Prism 5 software (Graph Pad, San Diego, CA, USA)를 이용하여 수행되었다. Western blot 결과와 ELISA 결과를 분석하기 위하여 일원변량분석(one-way ANOVA)을 이용하였다. Western blot 결과로 추정된 지방조직에서의 fetuin A 단백질 발현과 ELISA 결과로 추정된 혈장 순환 수준에서의 fetuin A 농도 사이의 상관관계를 확인하기 위하여 이변량 상관분석을 실시하고 피어슨의 상관계수(Pearson’s correlation coefficient)를 산출하였다. 체중변화와 OGTT 결과는 이원반복분산분석(two-way repeated measures ANOVA)에 의해 분석되었다. 일원변량분석과 이원반복분산분석 모두 Bonferroni 방법으로 모든 그룹 간에 사후검증하였다. 모든 통계적 유의수준은 p<.05로 설정하였다.
연구 결과
1. 체중증가, 식이섭취 그리고 식이효율
일반식이를 섭취한 집단과 고지방식이를 섭취한 집단 사이의 10주간의 체중 변화를 나타내었고 신체적 특성과 식이경향을 비교하였다(Table 1, Fig. 2). 그룹 간에 초기 체중에서는 유의한 차이가 없었고 최종 체중과 총 체중증가량 평균체중증가량 그리고 식이효율에서 모두 고지방식이를 섭취한 집단에서 유의하게 높게 나타났다[p<.001, ND-fed {normal diet-fed animals (n=14)} vs. HFD-fed {high-fat diet-fed animals (n=14)}]. 식이섭취량은 일반식이를 섭취한 동물들이 유의하게 많았다(p <.001). 본 연구의 일회성 운동이라는 중재 특성상 중재 및 희생 직전에 세부그룹의 분류가 이루어져 종단적 분석법에 근거한 체중증가, 식이섭취 그리고 식이효율에 미치는 영향을 비교하는 것은 통계적 의미성을 상실한다. 그러나 각 그룹의 개체 특성은 Table 1을 통해 확인할 수 있다.
2. 혈당 측정 결과
고지방식이를 섭취하여 비만이 유도된 실험동물은 일반식이를 섭취한 집단에 비해 전체적으로 AUC가 유의하게 넓었다(p <.001, NDfed vs. HFD-fed) (Fig. 3). 또한 20% 포도당 수용액을 경구투여한 후 30분 후에 유의하게 혈당이 높았고(p<.01, ND-fed vs. HFD-fed), 30분 후에는 유의한 차이가 없었다가 다시 60분과 120분에서 유의하게 혈당이 높게 측정되었다(p<.001, ND-fed vs. HFD-fed).
3. 지방조직에서의 fetuin A 단백질 발현
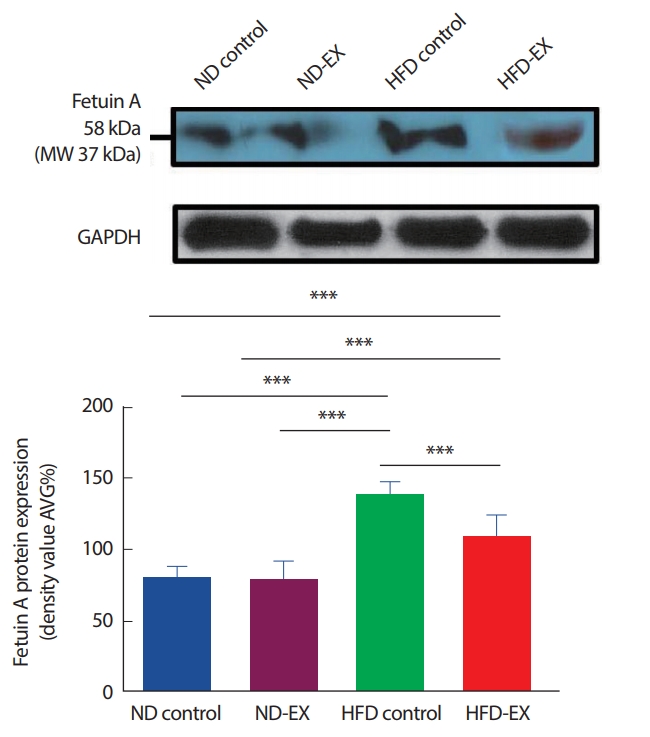
Western blot 결과 밴드의 픽셀 강도에 따라 fetuin A 단백질 발현량이 결정되었다(Fig. 4). HFD control은 모든 그룹에 비해 유의하게 발현이 높았다(p<.001). HFD control에 비해 HFD-EX에서 발현이 낮았다(p<.001). 그러나 HFD-EX는 ND control과 ND-EX군에 비해서는 유의하게 높았다(p<.001).
4. 혈장에서의 fetuin A 단백질 농도
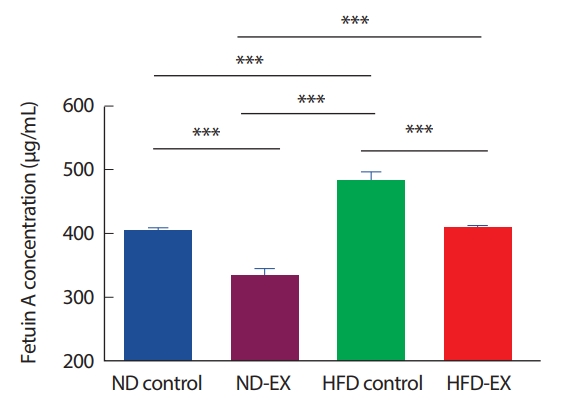
혈장에서의 fetuina A의 농도는 ND control에 비해 HFD control에서 유의하게 높았다(p <.001). ND-EX군은 모든 그룹과 각각 비교하여 유의하게 낮았다(p<.001 vs. ND control, HFD control, and HFD-EX). HFDEX군은 ND control군과는 차이가 없었고, HFD control에 비해 fetuin A 농도가 낮았으나(p<.001), ND-EX군보다는 높았다(p<.001)(Fig. 5).
논 의
본 연구에서 비만 유도 과정은 고지방식이 동물의 체중 및 인슐린저항성의 증가로 미루어 봤을 때 매우 효과적으로 이루어진 것으로 보여진다. 실험동물 희생 전에 체중이나 인슐린저항성은 측정되지 못하였다. 하지만 일회성의 운동만으로도 에너지 소모 및 탈수에 의해 체중이 감소할 가능성이 있고 인슐린 감수성 또한 증가할 수 있다는 충분한 근거들이 있다[35,36].
궁극적으로 본 연구자가 확인하고자 하였던 지방조직에서의 fetuin A는 장기간의 고지방식이로 인하여 HFD에서 ND에 비해 유의하게 증가한 것을 확인할 수 있었다. 비록 HFD-EX가 ND control과 ND-EX에 비해 유의하게 높았지만, HFD군에 비해 유의하게 발현이 낮아 비만이 유도된 동물에서는 일회성의 운동에 의해서 fetuin A가 감소하였음을 보여주었다. 한 가지 주목해야 할 부분은 ND control과 ND-EX 사이에는 차이가 없었다는 사실이다. 이러한 결과는 fetuin A의 발현은 지방조직 내의 세포내 섭취(endocytosis)와 성장에 관여하므로 비만상태가 아니더라도 일어나고 있으며 정상상태에서의 운동의 역할은 fetuin A의 감소에 영향을 미치지 못하는 것을 보여준다[37]. 더 나아가 fetuin A의 증가에는 비만으로 인한 과도한 에너지 유입이나 염증 그리고 인슐린저항성 등의 영향이 운동에 의한 역할보다 크다는 것을 짐작할 수 있다. 또한 fetuin A 발현의 정상범위가 존재할 것으로도 예상할 수 있다. 기존의 연구에서 fetuin A의 감소에 의해 혈관석회화가 증가할 수 있다는 보고가 있어 이러한 fetuin A 기준치 이하의 감소에 의한 부정적인 효과를 예상할 수 있다[38,39]. 혈장에서의 fetuin A의 농도는 ND-EX와 HFD-EX 모두 각각의 대조군인 ND control과 HFD control보다 유의하게 낮았기 때문에 운동에 의해 혈장 수준에서의 fetuin A는 감소하는 것으로 보여지며 이는 아마도 간에서 분비되므로 일회성 운동에 의한 효과는 지방조직보다 혈장에서 더욱 크게 나타나는 것으로 보여진다. 그러나 중요한 것은 비만동물을 대상으로 일회성의 운동을 실시하였을 때에는 지방조직과 혈장 모두에서 fetuin A가 감소하였다는 사실이다.
현재까지 혈장이나 혈청(serum)에서의 fetuin A 농도와 제2형 당뇨병 발병과의 양적인 상관관계가 잘 알려져 있기 때문에 제2형 당뇨병을 장기적인 관점에서 관리하기 위해서 운동은 더없이 효과적인 방법임을 본 연구에서 보여주고 있다. 기존의 연구에서 비당뇨병 여성을 대상으로 6주간의 유산소 운동을 실시하였을 때 fetuin A가 감소했다는 결과가 있었고[25], 당뇨병 환자를 대상으로 12주간의 유산소 운동을 실시하였을 때 혈청 fetuin A가 감소하였다는 보고가 있었다[40]. 이러한 사실은 제2형 당뇨의 예방뿐만 아니라 관리에도 운동이 매우 효과적으로 작용하며 그 중심에는 fetuin A가 있다는 것을 의미한다. 하지만 앞서 언급한 연구 중 당뇨병 환자를 대상으로 한 연구는 불과 몇 달전에 논문이 게재되었음에도 불구하고 여전히 지방조직에서 fetuin A의 발현은 측정되지 못하였다. 그리고 앞서 언급한 운동의 장기적인 효과뿐 아니라 본 연구와 같은 일회성의 운동만으로도 혈장 fetuin A의 농도를 낮추었다는 연구결과가 있었다[41]. 이러한 사실은 장기간의 운동과 일회성의 운동 모두 순환 수준에서의 fetuin A를 낮출 수 있다는 것을 뜻하며 본 연구의 결과와 일치하는 것으로 보인다.
일회성의 운동으로 인한 혈장 수준에서의 fetuin A의 감소는 여러 연구들에 의해 충분히 예상할 수 있는 결과였지만 일회성 운동에 의해 지방조직에서 fetuin A의 감소를 확인한 연구는 전무하였다. 따라서 본 연구에서 일회성 운동에 의해 감소하는 지방조직에서의 fetuin A의 발현은 처음으로 보고되는 결과이다. 일회성의 운동만으로도 비만인 상태의 지방조직에서 fetuin A가 감소한다는 것은 운동에 의한 비만의 개선에 따른 것이 아닌 운동에 의한 일시적인 반응이기 때문에 중요한 의미를 가진다. 일시적인 운동량의 변화와 함께 fetuin A가 변화하였다는 것은 최소한 장기간의 운동과 식이에 의한 비만의 개선과는 관련이 없는 것이기 때문에 운동에 의한 fetuin A가 아마도 비만의 개선 때문은 아니며 운동의 직접적 효과라는 것을 확실하게 보여주는 결과이기 때문이다. 하지만 연구결과의 해석에 유의해야 할 것은 본 연구에서의 fetuin A의 변화는 지방조직에서의 운동에 의해 수시간 이내에 일어나는 즉각적인 인슐린감수성의 증가와 면역반응의 변화와 함께 일어나는 변화이기 때문에 장시간 지속될 수 있는 효과가 아닐 가능성이 훨씬 높다는 점이다[42]. 따라서 fetuin A의 감소로 인한 일시적인 에너지 항상성의 정상화를 기대할 수는 있겠지만 장기간의 규칙적인 운동이 동반되지 않는다면 에너지 센싱 결함의 회복은 기대할 수 없을 것이다. 결론적으로 본 연구에서의 결과는 단지 일회성의 운동만으로도 fetuin A이라는 독립변인의 역할을 조절할 수 있다는 것 이상도 이하도 아니다. 따라서 장기적으로 fetuin A를 조절하여 지방조직에서의 에너지항상성을 유지하기 위해서는 장기간의 규칙적인 운동과 식이조절 등의 꾸준한 관리가 동반되어야 할 것이다.
본 연구에서 운동에 의한 비만 쥐의 지방조직에서 fetuin A 발현 감소가 최초로 확인되었다. 이전 연구에서는 혈장수준에서 많은 연구가 이루어졌지만 fetuin A는 지방조직에서도 특이적으로 발현되기 때문에 분명히 특정 자극에 의한 반응에는 혈장과 지방조직에서 각기 다른 반응을 보일 것으로 예상된다. 하지만 비만이나 운동에 의한 반응을 확인하여 혈장 수준과 지방조직에서의 fetuin A의 발현이 일률적으로 같다면 지방조직에서의 에너지항상성이 비정상적으로 조절되고 있다는 것을 혈장 수준에서의 fetuin A 측정만으로도 충분히 추측 가능할 것이다. 그렇기 때문에 우리는 fetuin A의 지방조직에서 단백질 발현과 혈장에서의 fetuin A 농도와의 상관관계 분석을 실시하여 본 연구에서 지방조직에서 분석했던 결과가 과연 이전의 혈장수준 연구들과 차별성을 가질 수 있는지 확인하고자 하였다.
전체 개체를 대상으로 상관관계를 분석하였을 때는 fetuin A가 비만과 인슐린저항성과 밀접한 관련이 있기 때문에 유의한 상관관계가 있는 것으로 나타났다(r=.709, p<0.01). 하지만 각 그룹 간에 경향 또한 일치하여야 fetuin A의 지방조직에서의 발현과 혈장 농도와의 관계가 더욱 밀접하다고 볼 수 있기 때문에 각 그룹 간에 따로 상관관계 분석을 추가로 실시하였다. 결과적으로 ND control과 ND-EX 그리고 HFD control 모두 상관관계가 없는 것으로 나타났고, HFD-EX가 유일하게 유의한 상관관계가 있었다(r=.885, p<.001). Fetuin A의 과발현은 비만으로 인한 지방조직의 과도한 지질축적에 의한 것으로 보여진다. 혈장 fetuin A 또한 간의 과도한 지방축적에 의해 합성이 증가한 것으로 보여지며 fetuin A의 지방조직에서의 발현과 혈장 농도 모두 일회성 운동에 의해 감소하였다. 이러한 사실은 고지방식이에 의한 비만은 간과 근육과 같은 우리 몸의 여러 장기에 지질축적을 증가시키고 여러 대사 이상을 초래할 수 있으며 이는 운동에 의해 개선될 수 있다는 기존의 연구에 근거하여 매우 자연스러운 결과로 보인다[43,44]. 따라서 지방조직과 혈장 농도 간에는 분명 비만으로 인해 증가하고 비만의 개선과 운동을 통한 개선에 의해 감소할 수 있다는 공통점이 있기 때문에 모든 개체를 포함하여 상관관계를 분석하였을 때에는 유의한 양의 상관관계를 나타낸 것으로 보인다.
다만 비만이나 인슐린저항성을 동반하지 않은 ND control과 ND-EX과의 상관관계가 어떤 의미를 가질 것인지에 대해서는 추가적인 논의가 필요할 것이다. 하지만 분명한 것은 이전의 연구에서 정상 쥐를 대상으로도 운동에 의해 fetuin A가 감소하였다는 결과가 있으므로 ND-EX에서 지방조직에서와 혈장에서의 모두 운동에 의해 fetuin A가 감소하였다면 양의 상관관계를 보였을 것이지만 관계가 없었고 오히려 추세선이 음의 기울기로 나타났다는 것은 정상수준에서의 지방조직과 혈액수준의 운동에 의한 반응에 차이가 있다는 것을 의미한다[29]. 또한 HFD control (Fig. 6D)에서 상관관계가 없었고 HFD-EX (Fig. 6E)에서 양의 상관관계가 나타난 것은 비만임에도 불구하고 지방조직에서의 fetuin A와 혈장에서의 fetuin A에 차이가 있을 수 있음을 의미함과 동시에 운동에 의한 fetuin A의 변화 경향은 비만인 경우 두드러지게 운동에 의한 반응이 지방조직과 혈장수준 모두에서 나타남을 의미하고 있다. 이러한 사실이 시사하는 바는 결국 간에서 합성되어 혈액으로 유리되는 fetuin A와 지방조직에서의 fetuin A의 반응이 다르게 나타나는 것을 의미한다. 또한 fetuin A가 간에서 합성되고 분비되어 혈장에 유리되어 작용하는 시간보다 지방조직에서 발현되는 반응이 좀 더 즉각적인 것으로 사료된다. 결론적으로 혈장 농도의 측정만으로는 지방조직에서의 fetuin A를 대변할 수 없다는 것을 의미하며 서론에 언급하였던 사람을 대상으로 하여 운동중재를 실시한 총 7건의 연구 중 혈장에서의 fetuin A의 발현이 효과가 없었던 3건의 연구에서는 아마도 혈장 fetuin A의 운동에 의한 반응이 지방조직에 비해서는 즉각적이지 않고 좀 더 장기적으로 조절되는 부분임을 충분히 고려하지 못하였기 때문일 수도 있을 것으로 생각된다[23,25,27]. 그러므로 앞으로는 비만과 fetuin A 관련 연구에서는 혈장수준에서만이 아니라 지방조직에서도 분석이 이루어져야 할 것이며, 스포츠과학 분야에서도 다양한 운동의 조건에 따른 지방조직에서의 fetuin A의 발현에 관한 연구가 비만인이나 동물모델을 대상으로 앞으로도 활발히 이루어져야 할 것이라고 생각된다.