서 론
적절한 골격근량(muscle mass)을 유지하는 것은 대사성 증후군[1], 당뇨[2], 노인성 근감소증[3] 등을 예방하여, 정상적인 대사와 독립적인 신체활동을 가능하게 한다. 또한 골격근량의 증가는 근력(strength)을 증가시켜 운동능력(performance)을 향상시키는 필수요소가 된다[4,5]. 골격근의 성장은 단백질 섭취와 같은 영양학적 요인과 신체활동에 의한 근수축 자극에 의해 단백질 합성 신호체계가 인산화 되거나 근 세포주기 조절 및 리모델링 요소들의 활성에 의해 조절된다. Serine/threonine kinase인 mammalian target of rapamycin (mTOR)을 중심으로 한 mTOR 신호체계는 mTOR의 하위 단백효소인 ribosomal protein S6 kinase (p70S6K)와 eukaryotic translation initiation factor 4E binding protein 1 (4E-BP1)을 인산화함으로써 단백질의 번역(translation)을 증가시킨다[6]. Extracellular signal-regulated kinase (ERK) 1/2, c-JUN NH2-terminal kinase (JNK), p38으로 구성된 mitogen-activated protein kinase(MAPK) 신호체계는 세포의 증식, 분화 등 세포주기를 조절하여 골격근의 성장을 조절하는 기전이다[7]. 특히, ERK1/2은 p90 ribosomal S6 kinase (p90RSK), eukaryotic initiation factor 4E (eIF4E) 등 myogenic 효소들을 활성 시킴으로써 골격근 세포에 직접적으로 영향을 미친다[8]. 근섬유의 근초(sarcolemma)와 기저막(basal lamina) 사이에는 골격근의 줄기세포인 위성세포(satellite cell)가 존재한다[9]. 이 위성세포는 평상시 대기상태(quiescent)로 존재하지만 운동과 같은 외적 자극에 의해 활성화되어 근세포의 필요에 따라 증식(proliferation)과 분화(differentiation)를 통하여 골격근의 재생 및 성장에 영향을 미친다[10].
현재까지 젊은 성인들의 골격근 비대(hypertorphy)를 위해 1 repetition maximum (RM)의 70-85% 강도로 저항성 운동을 수행하는 것이 권장되었다[11]. 그러나 고강도 운동을 수행하기 어려운 신체 허약자들에게 이 운동강도가 효과적인가에 대해서는 논란의 여지가 있으며, 최근 여러 선행연구들은 저강도 저항성 운동만으로도 고강도 운동과 유사한 골격근 단백질 합성을 보고하고 있다[12]. Burd et al. [13]은 젊은 성인 남성들을 대상으로 일회성 leg extension 운동을 90%RM의 고강도와 30%RM의 저강도로 더 이상 반복할 수 없을 때(set to failure)까지 실시한 결과 저강도 운동집단이 고강도 운동집단에 비해 유의하게 많은 총 운동량을 실시할 수 있었으며(90%RM, 710 ±30 kg; 30%RM, 1073±69.9 kg), 두 집단 간 대퇴사두근의 fractional synthesis rate (FSR)에 차이가 없음을 보고하였다. 또한 Mitchell et al. [14]은 젊은 성인남성들을 90%RM의 고강도와 30%RM의 저강도집단으로 나누어 주 3회씩 10주간 운동을 실시한 결과 두 집단 모두 대퇴사두근의 횡단면적이 증가하였으며, 두 집단 간에는 차이가 없음을 보고하였다. 저강도 운동수행이 고강도 운동과 유사한 골격근량의 증가가 나타난 연구들은 모두 세트마다 더 이상 운동을 수행할 수 없을 때까지 실시하는 volitional fatigue 방법을 적용하였다. 이 결과들은 골격근의 비대를 위해서는 고강도 저항성 운동(70-85%RM)만이 수행되어야 한다는 ACSM 권장 운동강도와는 다르게 저항성 운동의 방법에 따라 강도와 상관없이 근비대가 유도될 수 있음을 시사하고 있다. 또한 Williamson et al. [7]은 운동강도뿐만 아니라 골격근의 수축횟수에 의해서도 ERK1/2의 인산화가 조절될 수 있음을 주장함에 따라 운동강도 이외에 골격근량을 증가시키기 위한 다양한 운동방법의 접근을 고려해야 한다. 그러나 이 결과들을 뒷받침하기 위한 다양한 측면의 매커니즘 연구가 필요하며, 보다 정확한 운동처방적 접근을 위해 저강도 저항성 운동 중 어떠한 특성에 의해 운동의 강도와는 상관없이 근비대를 유도할 수 있는지 추가적인 연구가 필요하다.
따라서, 이 연구는 저강도 저항성 운동이 골격근의 재생과 성장 및 리모델링에 미치는 매커니즘을 확인하기 위해 일회성 운동 후 mTOR와 MAPK 신호체계의 인산화와 위성세포의 활성을 분석하였으며, 어떠한 운동특성이 저강도 저항성 운동에 의해 골격근의 비대를 유도하는 것인지 확인하기 위하여 운동강도, 근수축 횟수, 총 운동량의 차이에 따른 비교분석을 실시하였다.
연구 방법
1. 실험동물
이 연구에 사용된 실험동물은 유사한 형질을 위해 한 배 암컷과 수컷이 동종 교배된 12주령의 수컷 Sprague-Dawley rat이다. 실험진행 전 질병이나 이상증상을 보이는 실험동물은 대상에서 제외하였으며, 최종적으로 35마리를 7마리씩 5개의 집단으로 무선 배정하여 실험을 실시하였다. 동물실의 온도(22±2°C), 습도(50-60%) 및 명암조명(12/12시간; 명/암 주기)을 일정하게 유지하였으며, 동물윤리와 관련된 모든 절차는 한국체육대학교 동물실험윤리위원회로부터 승인받은 후 실시되었다(KNSU-IACUC-2017-04).
2. 실험설계 및 분석방법
1) 실험설계
실험집단은 일회성 저항성 운동 시 운동강도, 수축횟수, 총 운동량(운동강도×수축횟수)이 골격근의 단백질 합성 신호체계, 위성세포의 활성에 미치는 영향을 비교하기 위하여, 통제집단(Control group; Con, n=7), 저강도 저반복 집단(low-load, low-repetition; LLLR, n=7), 저강도 중반복 집단(low-load, medium-repetition; LLMR, n=7), 저강도 고반복 집단(Low-load, high-repetition; LLHR, n =7), 고강도 저반복 집단(High-load, low-repetition; HLLR, n =7)의 총 5집단을 설정하였다(Table 1). 이 집단을 통하여 동일한 운동강도 시 근수축 횟수가 미치는 영향(Con, LLLR, LLMR, LLHR 비교), 동일한 횟수의 근수축 시 운동강도에 의한 영향(Con, HLLR, LLLR 비교), 동일한 총 운동량 시 강도에 따른 영향(Con, HLLR, LLMR 비교), 저강도 운동이지만 고강도 운동보다 총 운동량이 많은 경우의 영향(Con, HLLR, LLHR 비교)을 비교함으로써 저강도 운동의 어떠한 운동적 특성이 단백질 합성 신호체계를 증가시킬 수 있는지 확인하였다.
2) 사다리 운동
이 연구를 위한 저항성 운동으로는 높이 1미터, 기울기 80도의 실험용 사다리를 이용하여 사다리 오르기 운동을 실시하였다. 실험을 실시하기 전 1주일간 주 3회의 사다리 오르기 사전 적응훈련을 실시하였으며, 사다리 운동에 충분히 적응된 후 본 운동 3일 전 Harris et al. [15]의 연구에 근거하여 각 개체의 1RM 측정을 실시하였다. 1RM 측정방법은 각 개체의 체중을 기준으로 최초 부하(50%체중)를 설정한 후 사다리 오르기를 성공할 경우 70%체중, 100%체중의 부하로 사다리 오르기를 실시하였으며, 이후 30 g씩 점증적으로 부하를 증가시켜 완전한 사다리 오르기가 가능한 마지막 무게를 1RM으로 설정하였다. 운동부하는 각 개체의 꼬리에 무게 추를 고정시키는 방법을 사용하였다. 이 운동은 측정된 1RM을 기준으로 각 개체 별 설정된 운동부하에 맞는 무게를 꼬리에 고정시켜 사다리의 바닥에서 상단까지 올라가는 것을 1회 반복(repetition)으로 설정하였다. 반복간 휴식시간은 없으며, 세트간 휴식은 2분으로 설정하였다.
3) 실험동물의 희생 및 표본준비
각 개체는 일회성 운동이 종료되는 시점부터 3시간의 안정기를 가진 뒤 실험동물을 희생하였다. 각 실험동물은 이산화탄소를 이용하여 마취시킨 뒤 사다리 오르기 운동 시 사용되는 주된 근육조직인 장모지굴근(flexor halluces longus, FHL)을 적출하였다[16]. 적출된 장모지굴근의 일부는 표현형 분석을 위해 O.C.T compound (Tissue-Tek; Sakura Finetek Europe, Zoeterwoude, The Netherlands)로 전 처리하였고, 일부는 단백질 발현 분석을 위해 액화질소에 급속냉동시켜 분석이 실시되기 전까지 -80°C에 보관하였다.
4) Western blot analysis
액화질소에 급속냉동된 FHL은 액화질소를 이용하여 잘게 부순 뒤 protease 와 phosphatase inhibitor cocktail이 첨가된 lysis buffer [25 mM Tris-Cl (ph 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethylsulfony1 fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 분주하여 4°C를 유지한 채 1시간 동안 화학적 분해과정을 거친 뒤 30분간 14,000 rpm의 속도로 원심분리하였다. 얻어진 상층액은 BCA protein assay 방법을 실시하여 단백질량을 측정한 후 4X SDS loading buffer (60mM tris pH6.8, 25% glycerol, 2% SDS, 14.4 mM 2-mercaptoethanol, 0.1% Bromophennol Blue)를 첨가하였다. 이와 같이 준비된 각 개체는 5-12% separating gel (30% acrylamide, 1.5M tris pH8.8, 10% Ammonium persulfate, TEMED)과 stacking gel (30% acrylamide, 1M tris pH6.8, 10% Ammonium persulfate, TEMED)에 분주하여 80 volt에서 전기영동을 실시하였다. 전기영동이 마쳐지면, Nitrocellulose Blotting Membrane(Life science, Germany)로 단백질의 이동을 위해 3M paper (Whatman)와 차례로 겹쳐 Mini trans blot을 장치한 뒤 Transfer buffer (190 mM glycine, 50 mM Tris-base, 0.05% SDS, 20% methanol)와 함께 80 volt로 90분간 전기영동을 실시하였다. Transfer가 완료되면 membrane을 5% BSA 용액(10 mM Tris-base, HCl-pH7.6, 0.5 M NaCl, 0.05% Tween20)에서 Blocking시킨 후 total Akt (9272s, Cell Signaling), phospho-AktSer473(9271s, Cell Signaling), total mTOR (2972s, Cell Signaling), phosphomTORSer2448(2971s, Cell Signaling), total p70S6K (9202s, Cell Signaling), phospho-p70S6KThr389 (9205s, Cell Signaling), total 4E-BP1 (9452s, Cell Signaling), phospho-4E-BP1Thr37/46 (9459s, Cell Signaling), total ERK (4695s, Cell Signaling), phospho-ERKThr202/Tyr204 (9101s, Cell Signaling), total JNK (9252s, Cell Signaling), phospho-JNKThr183/Tyr185(4671s, Cell Signaling), total p38 (9212s, Cell Signaling), phosphop38Thr180/Tyr182 (9211s, Cell Signaling), β-actin (sc-47778, Santa Cruz)의 1차 항체를 1:1,000의 농도로 blocking 용액과 혼합한 뒤 4°C에서 overnight하였다. 1차 항체 incubation이 완료되면 TBS-T 용액으로 10분 씩 3번 washing한 후 horseradish peroxidase (HRP) conjugated goat antirabbit(65-6120, ZYMED)와 anti-mouse (sc-2005, Santa Cruz)를 5% skim milk 용액과 1:5,000으로 혼합하여 90분간 incubation하였다. 2차 항체의 incubation이 끝나면, 최종적으로 TBS-T 용액을 이용하여 충분히 washing한 후 WBLR solution (Luminata Crescendo Western HRP Substrate, Millipole, USA)으로 단백질을 detecting 한 뒤 이미지 분석 시스템(Molecular Imager ChemiDoc XRS System, Bio-Rad, USA)으로 촬영하였다. 얻어진 이미지는 Quantity One 1-D Analysis Software (Bio-Rad, USA)를 이용하여 분석하였다.
5) Immunohistochemistry analysis
표현형 분석을 위해 O.C.T compound 처리된 조직은 Cryostat을 이용하여 5 μm 두께로 횡단절편을 낸 뒤 4% paraformaldehyde에 5분간 고정시켰다. 조직 고정이 마쳐지면 -20°C에 보관되어 있던 100% methanol에 10분간 담가둔 후 0.1% triton X-100과 혼합하여 만든 10% donkey serum (D9663, Sigma, USA)에 30분간 blocking하였다. 1차 항체는 안정기 위성세포 수를 확인하기 위해 Paired box 7 transcription factor(Pax7, MAB 1675, R&D systems, USA), 활성화된 위성세포를 확인하기 위해 Ki67 (ab16667, Abcam, England), 근막을 염색하기 위해 anti-laminin antibody (L9393, Sigma, USA)를 0.1M PBS에 1:100으로 혼합하여 4℃에서 overnight하였다. 1차 incubation 후 2차 항체로 Alexa flour 594 donkey anti-rabbit IgG antibody (MOP-A-21207, Life technologies, USA)와 Alexa flour 488 donkey anti-mouse IgG antibody (MOPA-21202, Life technologies, USA)를 0.1M PBS와 1:500으로 혼합하여 상온에서 4시간 동안 incubation하였다. 2차 항체의 incubation 후 4, 6-diamidino-2-phenylindole (DAPI)가 포함된 VECTASHIELD mounting medium (H-1200, VECTOR, USA)로 mounting하였다. 염색된 조직은 LEICA TCS SP8 confocal microscope (Leica Microsystem, Germany)을 이용하여 촬영하였다. 각 개체는 선행연구와 같이 신뢰도 있는 결과를 얻기 위해 OCT 처리된 조직을 3등분하여 각 부분에서 얻어진 절편으로부터 개체 당 200개 이상의 근섬유를 분석하였다[10].
연구 결과
1. 체중, FHL 무게 및 1RM
이 실험에 사용된 실험동물의 각 집단 별 체중, FHL 무게, 1RM은 Table 2와 같다. 본 운동 전 측정된 1RM은 집단 간 차이가 없었으며(p>.05), 일회성 운동 후 각 집단 간 체중, FHL의 무게도 차이가 없었다(p>.05).
2. 운동강도, 빈도 및 총 운동량
1RM 측정값에 따라 실시된 각 집단의 운동부하 및 총 운동량은 Table 3과 같다. 운동부하(무게 추)는 HLLR 집단이 30%RM의 저강도로 운동을 실시한 LLLR, LLMR, LLHR 집단에 비해 유의하게 높았다(p<.01, all). 실시된 총 운동량은 LLLR 집단이 LLMR, LLHR, HLLR 집단에 비해 유의하게 적은(p<.01, all) 반면, LLHR 집단은 LLLR, LLMR, HLLR 집단에 비해 높았다(p<.05, all).
3. Immunohistochemistry
1) 골격근 횡단면적, 안정기 위성세포, 근 핵
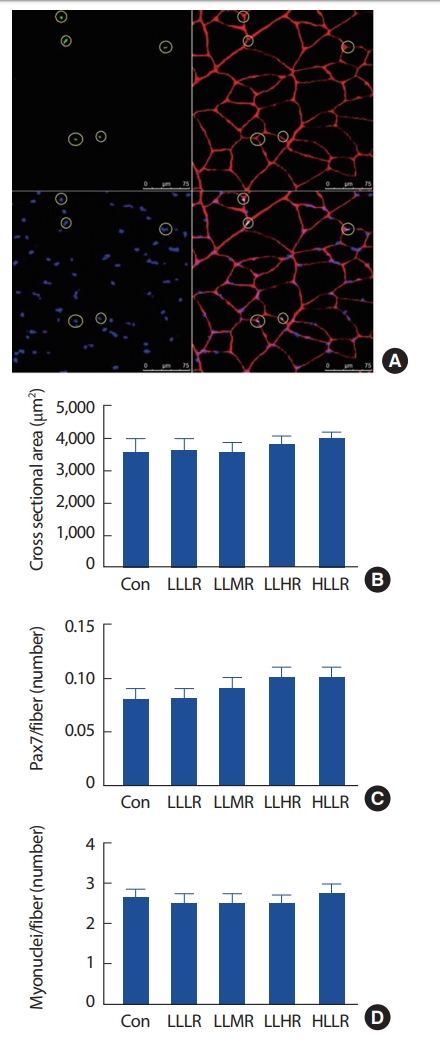
골격근의 횡단면적, 안정기 위성세포, 근 핵의 분석을 위해 Fig. 1A와 같은 이미지를 사용하였다. 일회성 운동 후 집단 간 FHL의 횡단면적(Fig. 1B, Con: 3563±397 μm2, LLLR: 3616±338 μm2, LLMR: 3554±294 μm2, LLHR: 3749±306 μm2, and HLLR: 3988±177 μm2), 위성세포의 수(Fig. 1C, Con: 0.08±0.01, LLLR: 0.08±0.01, LLMR: 0.09±0.01, LLHR: 0.10±0.01, and HLLR: 0.01±0.01) 및 근 핵의 수(Fig. 1D, Con: 2.66±0.18, LLLR: 2.49±0.24, LLMR: 2.50±0.22, LLHR: 2.47±0.24, and HLLR: 2.76±0.21)에는 차이가 없었다(p>.05, all).
2) 활성 위성세포(Active satellite cell)
활성 위성세포는 Fig. 2A와 같은 이미지를 사용하여, 총 위성세포 중 activation indicative 단백질인 ki67에 positive한 위성세포의 비율을 분석하였다. 일회성 운동 후 활성 위성세포의 비율은 Con과 LLLR 집단에 비해 LLHR (213%, p<.001 and 184%, p<.001, respectively)과 HLLR(225%, p<.001 and 194%, p<.001, respectively) 집단이 높았고, LLMR 집단에 비해 HLLR 집단(147%, p =.015)은 활성 위성세포 비율이 증가되었다(Fig. 2B).
4. Western blot
1) mTOR 신호체계
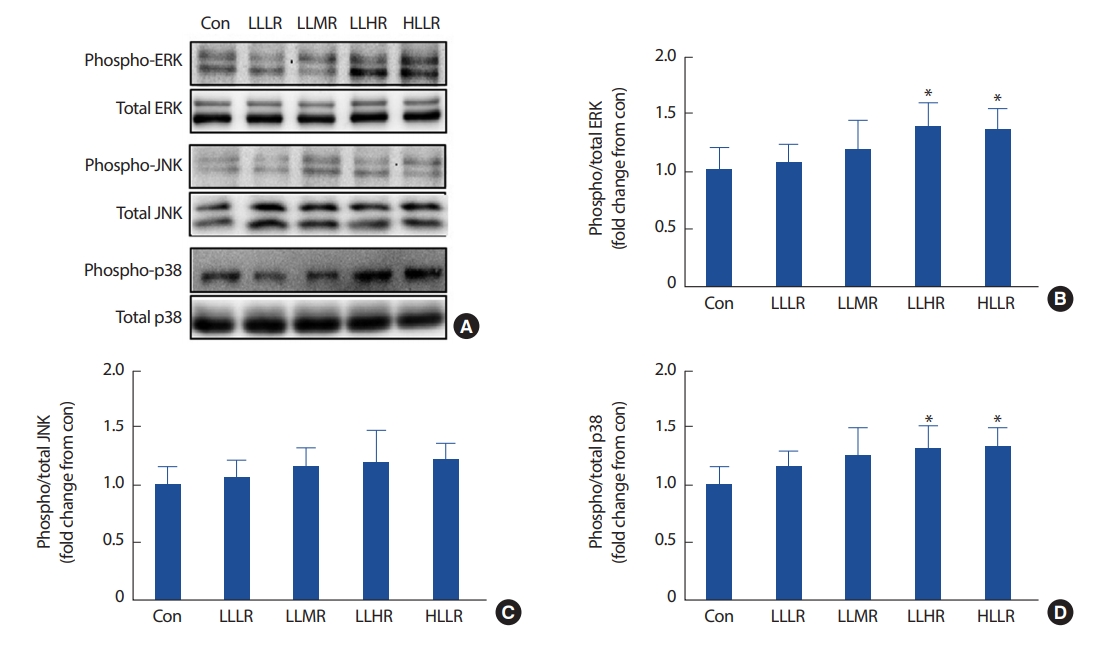
일회성 운동 후 Akt의 인산화는 각 집단 간 차이가 없었다(Fig. 3B, p >.05). 일회성 운동에 의한 mTOR의 인산화는 Con 집단에 비해 LLHR (137%, p =.038)과 HLLR (138%, p =.031) 집단이 유의하게 증가하였으며(Fig. 3C), p70S6K의 인산화도 Con 집단에 비해 LLHR (139%, p =.33)과 HLLR (142%, p =.16)집단이 유의하게 증가하였다(Fig. 3D). 4E-BP1의 인산화는 Con 집단에 비해 LLHR (135%, p =.008) 집단만이 유의하게 증가되었다(Fig. 3E).
논 의
최근 골격근 비대와 근력 향상을 위한 훈련방법으로서 저강도 고반복 저항성 운동의 효과가 전통적으로 실시되고 있는 고강도 저항성 운동과 유사함이 보고되고 있다[12]. 그러나 저강도 저항성 운동에 의한 대사적 매커니즘에 대한 연구는 많지 않다[17]. 이 연구의 주요결과는 일회성 고반복 운동을 실시한 LLHR 집단이 고강도 저항성 운동을 실시한 HLLR 집단과 유사하게 위성세포가 활성화되었으며, 단백질 합성을 조절하는 mTOR의 하위인자 p70S6K의 인산화와 세포의 주기를 조절하여 세포성장에 관여하는 ERK와 p38의 인산화를 증가시켰다. 특히, LLHR 집단은 일회성 운동 후 HLLR 집단과는 다르게 대조집단에 비해 증가된 4E-BP1의 인산화를 확인할 수 있었다. 이 결과들은 총운동량의 증가가 수반된 저강도 고반복 저항성 운동이 고강도 저반복 저항성 운동과 같이 골격근의 재생 및 성장을 위한 기전들을 활성 시킬 수 있음을 확인하였다.
위성세포는 골격근 내에 존재하는 줄기세포로 외적 자극에 의해 안정기 상태였던 위성세포가 활성화 된다. 활성화된 위성세포는 증식 단계를 거쳐 딸 세포 수를 증가시키고, 골격근의 요구에 따라 근아세포(myoblast)로 분화하여 골격근에 융합되거나 안정기 상태로 되돌아가는 자가재생(self-renewal)의 단계를 거치게 된다[18]. 이 연구에서 모든 운동집단은 일회성 운동 후 대조집단과 위성세포의 수에 차이가 없었지만(Fig. 1C), 저강도 고반복 운동을 실시한 LLHR 집단과 고강도 운동을 실시한 HLLR 집단은 대조집단에 비해 위성세포의 활성이 증가되었다(Fig. 2). 일회성 운동 후 위성세포의 자가재생을 확인하기 위해서는 운동 후 최소 24시간 이상의 충분한 시간이 요구되며, 운동 후 72시간에 가장 높은 위성세포 수의 증가를 확인할 수 있다[19]. 따라서 일회성 운동 후 3시간 시점에 골격근을 적출한 이 연구에서는 위성세포 수의 변화를 확인할 수 없었다. 그러나, LLHR 집단에서 활성화된 위성세포의 수가 증가함으로써 저강도 고반복 저항성 운동을 통해 고강도 운동과 유사한 위성세포의 자극이 가능함을 확인하였다. 이처럼 활성화된 위성세포는 분화되어 골격근에 융합됨으로써 직접적으로 골격근 대사에 참여하거나 자가재생에 의해 개체 수를 증가시켜 앞으로 일어날 골격근 비대나 재생의 잠재적인 요인(source)으로 작용된다[20]. 지금까지 선행연구들은 인간을 대상으로 위성세포가 골격근의 재생 뿐만 아니라 비대에도 필수적인 요소임을 보고하고 있다[21]. 그러나, McCarthy et al. [22]은 위성세포가 90% 이상 knock-out된 mice로 2주간 골격근수축을 유도하여 wild type mice와 동일한 근 비대를 보고함으로써 이 연구에 사용된 실험동물과 같은 설치류는 근 비대를 위해 위성세포가 필수적이지 않을 가능성을 제기하였다. 그러나 설치류에서 위성세포의 여부가 근 비대와 무관할지라도 위성세포는 유일하게 골격근에 새로운 근 핵을 증가시킴으로서 위성세포의 활성으로 인해 증가된 근 핵은 추후 근 비대 자극에 의한 단백질 합성 관련 전사인자(transcription)의 활발한 발현을 가능하게 한다[23]. 따라서 고강도 저항성 운동 뿐만 아니라 저강도 고반복 저항성 운동에 의해서도 위성세포의 활성을 통한 골격근의 재생 및 성장을 유도할 수 있다.
mTOR 신호체계는 단백질의 합성과 골격근량을 조절하는 주요 신호전달체계이다[24]. mTOR의 인산화는 그 하위인자인 p70S6K와 4E-BP1을 인산화 함으로써 리보솜에서 전사된 mRNA의 번역(translation)을 개시(initiation)하고 확장(elongation)하는데 주요한 역할을 담당한다[25]. 이 연구는 mTOR의 상위인자인 Akt의 인산화는 확인할 수 없었지만(Fig. 3B), LLHR과 HLLR 집단에서 대조집단에 비해 유의하게 증가된 mTOR, p70S6K의 인산화를 확인하였다(Fig. 3C, 3D). 일반적으로 mTOR는 insulin growth factor 1 (IGF1)이 IGF1 수용체와 결합됨으로 Akt를 인산화 시켜 하위인자인 mTOR를 활성 시킨다[26]. 그러나 운동과 같은 골격근의 수축은 Akt와는 독립적으로 mTOR를 인산화시킬 수 있으며[27], Ogasawara et al. [28]의 연구결과를 통해 일회성 운동 시 Akt의 인산화가 증가되었더라도 짧은 반감기를 고려한다면 운동 후 3시간이 지난 시점에서 자극이전의 상태로 회복되었을 가능성이 있다. LLHR 집단과 HLLR 집단은 모두 p70S6K의 인산화를 증가시켰다. 이러한 결과는 고강도 저항성 운동에 의해 p70S6K의 인산화가 증가된다는 여러 선행연구들과 일치할 뿐만 아니라[29] 저강도 고반복 저항성 운동에 의해서도 효과적으로 p70S6K의 인산화를 증가시킬 수 있었으며, HLLR 집단에서는 차이가 나지 않는 4E-BP1의 인산화가 저강도 고반복 저항성 운동에 의해 증가하였다(Fig. 3E). 4E-BP1은 p70S6K와 같이 mTOR의 downstream에 존재하고 있지만 운동방법, 영양섭취, 호르몬 등 자극의 종류에 따라 서로 다른 인산화 반응성을 지니고 있다[11, 30]. 따라서 4E-BP1 인산화가 LLHR 집단에서만 증가된 명확한 이유를 확인할 수는 없었지만, HLLR 집단에서 실시된 고강도 운동이 FOXO1 단백질의 발현을 증가시켜 4E-BP1의 mRNA 발현을 촉진함으로써 4E-BP1의 인산화를 억제하였을 가능성이 있다[11]. 이 연구결과들은 저강도 고반복 저항성 운동에 의해 효과적으로 단백질의 번역을 개시 및 확장함으로써 고강도 운동과 같은 근 비대를 유도하기에 충분한 효과가 있음을 시사한다.
MAPK는 myogenic 전사인자들이나 세포주기를 조절하여 골격근의 증식 및 분화에 작용하는 필수기전이다[8,31]. Williamson et al. [7]은 젊은 남성들을 대상으로 일회성 운동 실시한 후 Widegren et al. [32]의 연구결과와 비교분석을 통하여 운동강도뿐만 아니라 근수축 횟수가 증가함에 따라 ERK의 인산화가 증가할 가능성을 제기하였다. 그러나 이 연구결과는 고강도로 12번 사다리 오르기를 실시한 HLLR 집단은 ERK의 인산화가 대조집단에 비해 증가하였지만, 저강도로 동일한 횟수(12회)의 사다리 오르기를 실시한 LLLR 집단은 ERK의 인산화를 유도하지 못하였다(Fig. 4B). 반면, 저강도 고반복을 실시한 LLHR 집단은 HLLR 집단과 같이 효과적으로 ERK의 인산화를 증가시킬 수 있었다. 이 결과들을 통해 ERK의 인산화는 근수축 횟수에만 국한되지 않고 충분한 운동강도와 운동량에 따라 반응할 수 있음을 확인하였다. 또한 LLHR과 HLLR 집단의 JNK 인산화는 대조집단과 차이가 없었으며(Fig. 4C), 대조집단에 비해 증가된 p38의 인산화를 나타냈다(Fig. 4D). 이처럼, 저강도 고반복 저항성 운동은 전반적으로 고강도 저항성 운동과 유사한 형태의 MAPK 인산화를 유도하는 것을 확인할 수 있었다.
이 연구의 제한점으로 동일한 운동강도에서 골격근수축 횟수에 따른 효과를 검증 시 집단 간 총 운동량의 차이가 수반되었으며, 동일한 근수축 횟수 시 운동강도에 따른 차이를 검증할 경우에도 총 운동량의 차이가 수반될 수밖에 없었다. 따라서, 실시된 운동강도, 총 운동량에 따른 골격근 비대관련 신호전달기전들의 차이를 비교하기에는 효과적이었지만, 총 운동량을 동일하게 하면서 근수축 횟수나 운동강도에 의한 효과를 검증하기 위해서는 추가적인 연구설계가 요구된다.
종합해보면 저강도 저항성 운동은 운동강도와는 상관없이 증가된 총 운동량에 의해 위성세포의 활성과 단백질 합성기전들이 인산화됨을 확인할 수 있었다. 뿐만 아니라 4E-BP1의 인산화는 저강도 고반복 저항성 운동에 의해 고강도 운동보다 효과적으로 유도되었다. 이와 같이 저강도 저항성 운동에 의한 근 비대 관련 기전의 활성이 고강도 저항성 운동과 유사한 결과를 보인 것은 운동강도보다는 증가된 총 운동량에 근거한다.