


INTRODUCTION
Aging is a natural process that is associated with a decline in physiological and metabolic functions, leading to various age-related diseases. One of the hallmarks of aging is cellular senescence, a state of irreversible growth arrest that occurs in response to various stresses, including oxidative stress, DNA damage, and inflammation [1,2]. Vascular endothelial senescence is a critical contributor to age-related vascular dysfunction, which is characterized by impaired endothelium-dependent vasodilation and increased vascular inflammation [3]. Senescent endothelial cells can lead to reduced blood flow to tissues and organs, which can cause isch-emia and tissue damage. This can contribute to the development of conditions such as peripheral artery disease, coronary artery disease, and stroke [4]. Senescent endothelial cells can also contribute to the formation of blood clots by producing less nitric oxide, a molecule that helps regulate blood flow and prevent clotting. This can increase the risk of conditions such as deep vein thrombosis and pulmonary embolism [5].
Regular exercise has been shown to have anti-aging effects on the vascular system by improving endothelial function and diminishing oxidative stress and inflammation [6]. One of the primary mechanisms by which exercise prevents endothelial damage and senescence is through the activation of endothelial nitric oxide synthase (eNOS) [7]. eNOS is an enzyme that produces nitric oxide (NO), a potent vasodilator and anti-inflammatory molecule that plays a critical role in maintaining endothelial function [8]. Exercise-induced shear stress on the endothelium increases the production of NO by eNOS, leading to improved endothelial function and decreased oxidative stress and inflammation [9,10]. Exercise also promotes the production of antioxidants and anti-inflammato-ry molecules, which can protect against oxidative stress and inflammation-induced endothelial damage and senescence [11]. Exercise-induced increases in the production of superoxide dismutase, glutathione peroxi-dase, and catalase can enhance the cellular antioxidant defense system, while exercise-induced increases in the production of interleukin-10 and other anti-inflammatory cytokines can reduce inflammation and protect against endothelial damage [12]. However, adherence to an exercise regimen can be difficult for some individuals, particularly for those with limited mobility or other health conditions. Therefore, there is a growing interest in identifying natural compounds that mimic the beneficial effects of exercise on the cardiovascular system [13].
Resveratrol, a natural polyphenol found in grapes and red wine, has been suggested to have exercise-mimetic effects on the cardiovascular system, including improving endothelial function and reducing oxidative stress [14]. Resveratrol has been shown to activate a variety of signaling pathways involved in exercise-induced adaptations, including AMP-activated protein kinase (AMPK) and sirtuins. These signaling pathways regulate mitochondrial biogenesis, oxidative metabolism, and inflammation, which are important determinants of vascular function [15]. Also, resveratrol may improve cardiovascular health by increasing eNOS expression and/or activity in endothelial cells, which leads to increased production of nitric oxide and improved blood flow [16,17].
In recent years, there has been an increasing number of studies investigating the potential of resveratrol to prevent vascular endothelial senescence. These studies have provided evidence that resveratrol treatment can prevent cellular senescence in vascular endothelial cells by reducing oxidative stress and activating sirtuins [18].
The purpose of this study is to investigate the effects of resveratrol as an exercise-mimetic for the prevention and improvement of oxidative stress-induced cellular senescence in vascular endothelial cells.
METHODS
1. Cell culture
HUVECs from Lonza were cultured in M199 medium supplemented with 20% fetal bovine serum and endothelial cell growth supplement (Sigma-Aldrich no. E2759) at 37°C in a 5% CO2 atmosphere until pas-sages 3-5. Premature endothelial senescence was induced by treating HUVECs with 100 μM H2 O2 for 1 hour. After trypsinization and re-seeding at a density of 1.0×105 cells per well of a 6-well plate, cells were cultured in M199 complete media containing for 3 days. Following the 3 days of aging, HUVECs were exposed to 20 dyne/cm2 of orbital-flow shear stress (OFSS) for 72 hours in 3 mL cell culture medium. For shear experiments, HUVECs were exposed to the physiological levels of shear stress, 20 dyne/cm2 in orbital shaker. The intensity of OFSS was calculat-ed as τ max=α√(p×η(2πf)3) (α=orbital radius, p = medium density, η= iscosity of the medium, f=frequency of rotation). SIRT1 activation was induced by treating HUVECs with 20 or 50 µM concentration of resveratrol (trans-3,4’,-5-trihydroxystilebene) (Sigma, R5010) for 24 hours, while the solvent control dimethyl sulfoxide (DMSO) was used for the control. For SIRT1 inhibition, HUVECs were washed with growth medium and exposed to 60 µM concentrations of sirtinol (Calbiochem no. 566320), a cell-permeable 2-hydroxy-1-napthaldehyde derivative and a chemical inhibitor of NAD+-dependent protein deacetylases of sirtuins.
2. Senescence-associated β-galactosidase staining
The proportion of Senescence-Associated β-Galactosidase (SA-β-gal) positive cells was determined as described previously [19]. First, the cell growth medium was removed from the cells and they were washed with PBS. After that, the cells were fixed in a solution containing 20% formal-dehyde and 2% glutaraldehyde for 15 minutes. Next, the cells were incubated overnight in β-Galactosidase staining solution at 37°C in a dry incubator. This solution consists of 20 mg X-gal dissolved in 1 mL N-N-dimethylformamide (DMF), 400 mM citric acid/sodium phosphate (pH 6.0), 1.5 M NaCl, 20 mM MgCl2, 500 mM potassium ferrocyanide, and 500 mM potassium ferricyanide. The percentage of SA-β-gal positive cells was determined by counting the number of blue-stained cells out of the total number of cells in the same field of view at 200x total magnification.
3. Immunoblot
The antibodies used for the western blot were obtained from the fol-lowing sources: rabbit monoclonal anti-SIRT1 (Cell signaling no. 2496), rabbit monoclonal anti-p21 (Cell signaling no. 2947), and mouse monoclonal anti-α-Tubulin (Sigma no. T9026) for use as the internal control. Human umbilical vein endothelial cells (HUVECs) were homogenized in ice-cold RIPA lysis buffer with protease inhibitor (Roche no. 11836153001). Proteins from the cell lysates were separated using Tris-glycine SDS PAGE and transferred onto an Immobilon-P PVDF membrane for standard ECL western blotting. Chemiluminescence was used to detect total protein expression. Image J software from the National Institute of Health was used to perform band densitometry analyses and compare experimental sets. The values obtained from Image J are arbitrary units. The densities of the selected protein bands were expressed as a ratio to the densities of the internal control gene, α-Tubulin.
4. Matrigel tube formation assay
The Matrigel Basement Membrane Matrix (BD no. 356237) was thawed, and 50 μL of the Matrigel matrix was added to each well of a 96-well plate while keeping it on ice. The plate was then placed in a 37°C incubator for 30 minutes to allow the Matrigel to solidify. Next, 2.0×104 cells in 200 μL of complete media were added to the Matrigel-coated wells of the 96-well plate. Tube formation was observed under a micro-scope at 8, 12, and 24 hours after seeding, using a 100x magnification (Axiovert 40 CFL, Zeiss). The tube formation capacity of the endothelial cells was determined by counting the number of tubes formed.
5. Scratch assay
Cells were seeded and cultured in a 12-well plate with a starting cell density of 1.0×105 cells, and cultured until they reached over 90% con-fluence. A line was drawn on the bottom of the culture dish using a marker. The cells were then rinsed with PBS, and wounds were generat-ed by making scratches through the cells using a sterile 10 µL pipette tip moving perpendicular to the line that was drawn. After 16 hours, the size of the scratches was microscopically examined using a 100x magnification (Axiovert 40 CFL, Zeiss).
6. MitoSox staining
In each well of a 12-well plate, HUVECs were treated with 5 μM MitoSox reagent (Molecular Probes no. M36008) in 1 mL of HBSS containing CaCl2, MgCl2, and MgSO4. The cells were then incubated in a light-protected humidified 37°C incubator for 10 minutes. After washing the cells 3 times with HBSS, cell fluorescence was evaluated using in-verted fluorescence microscopy (IX81, Olympus).
7. Statistical analysis
Statistical analysis was performed by using IBM SPSS Statistics 27.0. The data is presented as mean±standard error of the mean (SEM). Dif-ferences between experimental conditions were analyzed by using analysis of variance followed by Fisher's least significant difference test for post-hoc testing. A statistical significance level of p <.05 was applied to all analyses.
RESULTS
1. Shear stress improves premature endothelial senescence
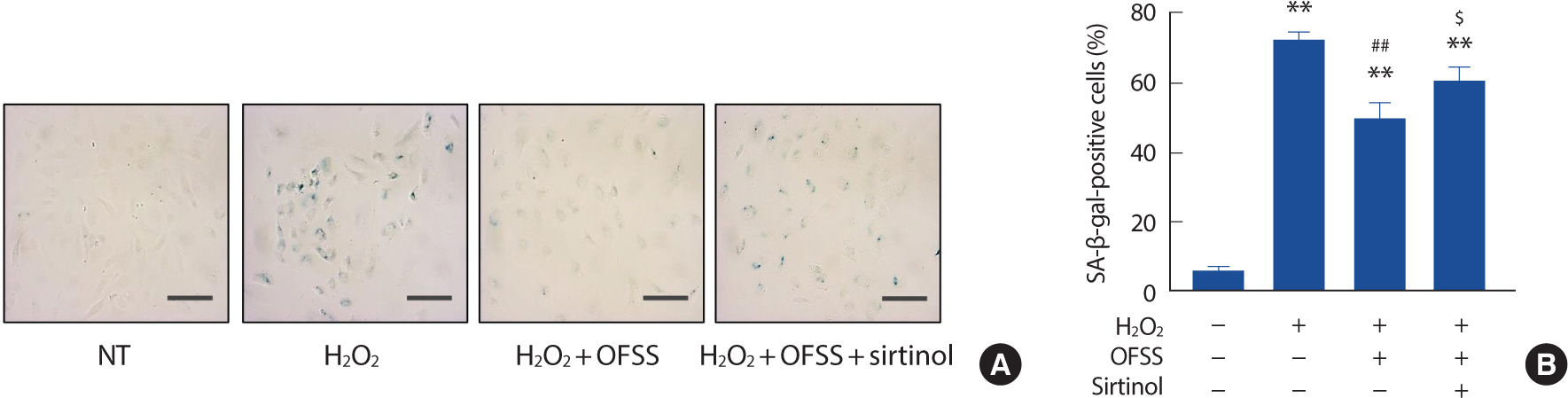
In order to explore how shear stress could reverse premature endothelial senescence, an orbital-flow shear stress (OFSS) was utilized on HUVECs that had been induced into senescence through exposure to H2 O2. The application of OFSS was found to have a partial restorative effect on the level of premature endothelial senescence. Nonetheless, the positive effect of shear stress was nullified when SIRT1 inhibition was introduced (Fig. 1A, B).
Fig. 1.
Fig. 1.Laminar shear stress normalizes premature endothelial senescence. Representative SA-β-gal staining pictures. Cell morphologies were microscopically examined at 200x magnification (A). Percentage of SA-β-gal-positive staining cells. The percentage of SA-β-gal positive cells was determined by counting the number of stained cells out of the total number of cells in the same field. Cells were counted in 90 independent fields (B). Scale bar=100 μm. NT, non-treated control; OFSS, orbital-flow shear stress.
** p<.01 compared to NT; ## p<.01 compared to H2 O2 treatment; $ p<.05 compared to H2 O2 +OFSS treatment.
2. Resveratrol prevents oxidative stress-induced premature endothelial senescence
In order to investigate how SIRT1 affects the inhibition of H2 O2-in-duced premature endothelial senescence, HUVECs were exposed to resveratrol, the SIRT1 activator, at concentrations of 20 or 50 μM for 24 hours prior to the application of 100 μM H2 O2. The H2 O2-treated cells were then cultured in normal cell-culture media for an additional 3 days. The percentage of SA-β-gal-positive cells, which indicates premature senescence, was significantly increased in the H2 O2-induced cells. However, this effect was partially reversed in cells that were pre-treated with resveratrol (Fig. 2A, B). The results of western blotting analysis of p21 expression were consistent with the SA-β-gal staining results (Fig. 2C). Also, the expression of SIRT1 was preserved in HUVECs that were pre-treated with resveratrol (Fig. 2D).
Fig. 2.
Fig. 2.Resveratrol prevents H2 O2-induced premature endothelial senescence. Representative SA-β-gal staining pictures. Cell morphologies were microscopically examined at 200x magnification (A). Percentage of SA-β-gal-positive staining cells. Percentage of SA-β-gal positive cells was determined by counting the number of stained cells out of the total number of cells in the same field. Cells were counted in 90 independent fields (B). p21 gene expression analyzed by Western blotting (C). SIRT1 gene expression analyzed by Western blotting. α-Tubulin was used as a loading control (D). The bar graphs are results of densitometry analyses. Each column represents mean±SEM from 3 independent experiments. Scale bar=100 μm.
NT, non-treated control; RSV, resveratrol.
* p<.05 and ** p<.01 compared to NT; # p<.05 compared to H2 O2 treatment.

3. Resveratrol attenuates ROS production in senescent endothelial cells
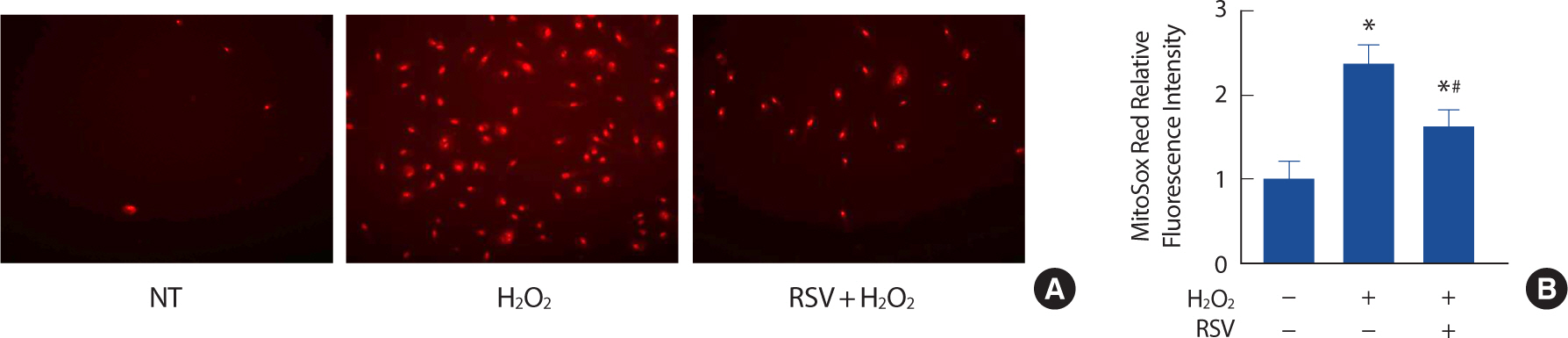
In order to gain a better understanding of how SIRT1 exerts its anti-oxidative effects, we examined the levels of ROS production in endothelial cells that were pre-treated with 20 μM resveratrol. Resveratrol, which is a known activator of SIRT1, was found to significantly decrease the levels of ROS production in endothelial cells that were induced to senescence by the H2 O2 treatment and aging (Fig. 3A, B).
Fig. 3.
Fig. 3.Resveratrol attenuates ROS production in H2 O2-induced senescent endothelial cells. Representative pictures of MitoSox staining (A). Relative fluorescence intensity in different conditions (B). Each column represents mean±SEM from 3 independent experiments. NT, non-treated control; RSV, resveratrol.
* p<.05 compared to NT;.# p<.05 compared to H2 O2.
4. Resveratrol prevents oxidative stress-induced endothelial dysfunction
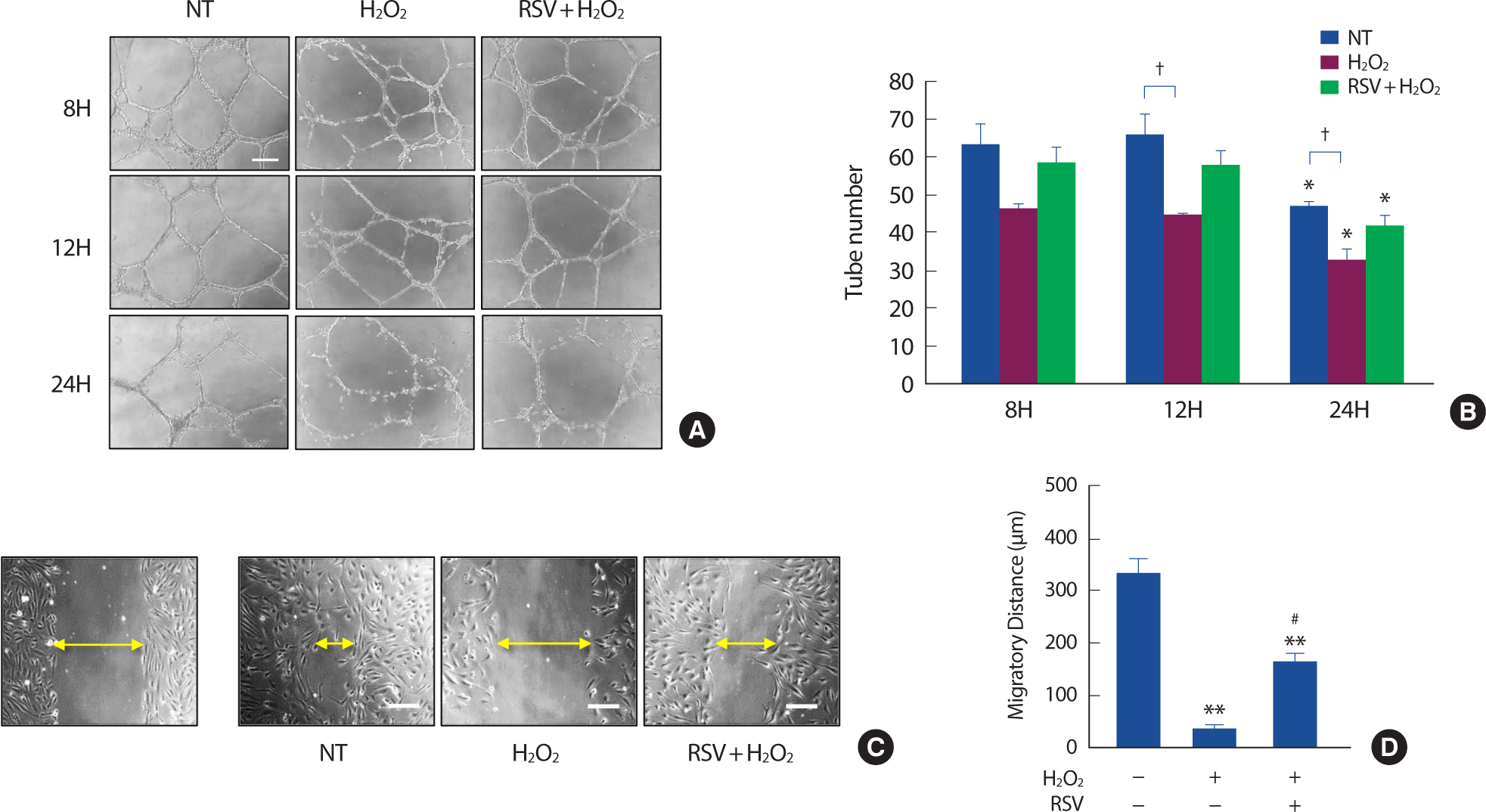
In order to investigate the impact of cellular senescence on endothelial function and the role of SIRT1 in preventing endothelial dysfunction, we conducted the Matrigel tube formation assay to assess a vasculogenic capacity and the cell scratch assay to assess cell migration capacity. We found that the tube numbers (Fig. 4A, B) and the migratory distance (Fig. 4C, D) of HUVECs that were treated with H2 O2 were significantly decreased. However, the ability to form tubes (Fig. 4A, B) and migrate (Fig. 4C, D) was partially maintained when SIRT1 was activated by the pretreatment of 20 μM resveratrol.
Fig. 4.
Fig. 4.Resveratrol prevents H2 O2-induced endothelial dysfunction. Representative pictures of Matrigel tube formation assay (A). Tube numbers in different conditions (B). Representative pictures of cell scratch assay. Left, HUVECs immediately after making the scratch. Right, Scratch size 16h after scratch (C). Cell migratory distance in different conditions 16h after scratch (D). Each column represents mean±SEM from 3 independent experiments. NT, non-treated control; RSV, resveratrol. Scale bar=200 μm.
* p<.05 and ** p<.01 compared to NT; # p<.05 compared to H2 O2 treatment; † p<.05 compared between groups.
DISCUSSION
The present study aimed to investigate the aerobic exercise-mimetic effects of resveratrol on the prevention of vascular endothelial senescence. The results of the study indicated that resveratrol can prevent cellular senescence in vascular endothelial cells by reducing oxidative stress and activating sirtuins. The findings of this study add to the growing body of evidence suggesting that resveratrol may have exercise-mimetic effects on the cardiovascular system.
Aging is a natural process that is associated with a decline in physiological and metabolic functions, leading to various age-related diseases [2]. One of the hallmarks of aging is cellular senescence, a state of irreversible growth arrest that occurs in response to various stresses, including oxidative stress, DNA damage, and inflammation [1]. Vascular endothelial senescence is a critical contributor to age-related vascular dysfunction, which is characterized by impaired endothelium-dependent vasodilation and increased vascular inflammation. Regular exercise has been shown to have anti-aging effects on the vascular system by improving endothelial function and diminishing oxidative stress and inflammation [7]. However, adherence to an exercise regimen can be difficult for some individuals, particularly for those with limited mobility or other health conditions [20]. Therefore, there is a growing interest in identifying natural compounds that mimic the beneficial effects of exercise on the cardiovascular system.
Resveratrol, a natural polyphenol found in grapes and red wine, has been suggested to have exercise-mimetic effects on the cardiovascular system, including improving endothelial function and reducing oxidative stress [21,22]. Resveratrol has been shown to activate a variety of signaling pathways involved in exercise-induced adaptations, including AMP-activated protein kinase and sirtuins. These signaling pathways regulate mitochondrial biogenesis, oxidative metabolism, and inflammation, which are important determinants of vascular function [23]. In recent years, there has been an increasing number of studies investigating the potential of resveratrol to prevent vascular endothelial senescence [24]. These studies have provided evidence that resveratrol treatment can prevent cellular senescence in vascular endothelial cells by reducing oxidative stress and activating sirtuins [25].
Our results show that resveratrol treatment can prevent cellular senescence in vascular endothelial cells induced by oxidative stress. Specifical-ly, we found that resveratrol treatment led to a decrease in the proportion of senescence-associated β-galactosidase positive cells, a well-established marker of cellular senescence. This finding is consistent with previous studies that have demonstrated the anti-aging effects of resveratrol on the cardiovascular system, including its ability to improve endothelial function and reduce oxidative stress and inflammation [26,27].
Furthermore, the current results also showed that the anti-senescence effects of resveratrol were mediated, at least in part, by the activation of SIRT1, a key regulator of cellular senescence, metabolism, and oxidative stress. Treatment with resveratrol increased the expression of SIRT1, while inhibition of SIRT1 with sirtinol blocked the anti-senescence effects of resveratrol. These findings suggest that the activation of SIRT1 by resveratrol may play an important role in preventing vascular endothelial senescence. And these findings are consistent with previous studies showing that resveratrol treatment can activate SIRT1 and prevent cellular senescence in various cell types [25,28,29].
One limitation of our study is that we used a cell culture model of premature endothelial senescence induced by hydrogen peroxide treatment. While this model has been widely used to study the mechanisms of cellular senescence, it does not fully recapitulate the complex patho-physiology of age-related vascular dysfunction in vivo. Therefore, future studies are needed to validate our findings in animal models and in human clinical trials.
The current study provides further evidence for the anti-aging effects of resveratrol on the vascular system and highlights its potential as an exercise-mimetic compound for the prevention of age-related vascular dysfunction. The activation of SIRT1 by resveratrol appears to be a key mechanism underlying its anti-senescence effects, suggesting that targeting SIRT1 may be a promising strategy for the prevention and treatment of age-related vascular diseases. Further research is needed to elucidate the precise mechanisms by which resveratrol and other exercise-mimetic compounds exert their beneficial effects on the cardiovascular system and to determine their safety and efficacy in human populations.
CONCLUSION
The results of this study demonstrate that laminar shear stress can mitigate the effects of oxidative stress-induced premature endothelial senescence, and that SIRT1, a protein involved in cellular repair processes, is crucial in this anti-senescence response. Resveratrol, a compound known to activate SIRT1, may be a potential substitute for exercise in preventing age-related damage to the endothelium by increasing resistance to oxidative stress and improving endothelial function.


