서 론
파킨슨질환(Parkinson’s Disease, PD)은 알츠하이머질환(Alzheimer’s disease) 다음으로 환자 수가 많은 신경퇴행성질환으로 운동완서(bradykinesia), 자세반사이상(postural reflex) 및 강직(rigidity) 등의 운동성 장애가 나타난다[1,2]. 현재까지 파킨슨질환에 대한 정확한 발병기전과 치료책은 알려져 있지 않지만, 중뇌 흑질(substantia nigra, SN)에 존재하는 도파민 신경세포의 감소와 비정상적인 단백질의 응집체인 루이소체(lewy bodies, LBs)가 병리학적 특징으로 제시되고 있다[3]. 이중 루이소체의 주성분으로 알려진 α-synuclein (α-syn) 단백질은 주로 시냅스 전 종말에서 신경전달물질을 방출하는 역할을 담당하지만 비정상적으로 과도하게 축적되면 산화적 스트레스를 증가시켜 도파민 신경세포의 소실과 함께 운동기능의 장애를 일으킨다[4,5]. 또한, α-syn 단백질의 축적은 mtDNA 손상, 단백질 분해 기능저하, 미토콘드리아 관련 효소들의 활성을 감소시켜 미토콘드리아 기능이상을 일으키는 것으로 보고되었다[6-10].
미토콘드리아(mitochondria)는 ATP 생산, 칼슘의 항상성, 세포의 생존 및 성장에 매우 중요한 세포소기관이다. 특히, 신경세포의 미토콘드리아는 신경전달물질을 분비하기 위해 필요한 에너지를 공급하여 신경세포의 성장과 기능을 조절한다[11]. 일반적으로 미토콘드리아의 구조는 미토콘드리아 융합(fusion)과 분열(fission)에 의해 결정되며, 이러한 반복적인 과정을 통해 세포의 항상성이 조절되지만 이러한 역동성의 균형이 무너지면 산화적 스트레스를 일으키고 결국 병리학적 과정으로 진행되는 것으로 보고되었다[11]. 따라서 미토콘드리아의 구조와 기능을 유지하기 위해 미토콘드리아 역동성(mitochondria dynamic)을 정상적으로 유지하는 것이 신경세포의 생존과 성장에 중요하다고 볼 수 있다.
하지만 최근 α-syn 단백질의 축적은 미토콘드리아 역동성의 장애를 일으켜 미토콘드리의 구조와 기능을 저하시킨다고 제시되고 있는 가운데, Xie et al. [12]과 Kamp et al. [13]은 과 발현된 α-syn 단백질이 미토콘드리아 융합을 감소시키고 분열을 증가시켜 미토콘드리아 분절과 함께 미토콘드리아 기능이상을 일으키는 것으로 보고하였다. 또한, α-syn의 축적은 미토콘드리아 분열을 일으키는 dynamin related protein 1 (Drp 1) 단백질의 활성을 증가시켜 미토콘드리아 구조 이상과 신경세포의 사멸을 일으킨다고 보고되었다[14,15]. 따라서 α-syn 단백질을 감소시키는 것은 미토콘드리아 역동성을 정상적으로 유지하여 미토콘드리아 구조와 기능을 개선시킬 수 있는 효과적인 방법이라고 생각된다.
최근 신체활동은 신경퇴행성 질환과 관련되어 신경재생, 신경세포 생존, 행동장애, 인지기능 및 미토콘드리아 기능 개선에 효과적인 방법으로 제시되고 있다[16-18]. 특히, 파킨슨질환을 대상으로 트레드밀 운동은 tyrosine hydroxylase (TH)의 활성을 증가시켜 도파민 신경세포의 소실을 억제하고[19], 신경성장인자인 brain-derived neurotrophic factor (BDNF)와 신경가소성을 회복시켜[20,21] 운동기능의 장애가 개선된 것으로 보고되었다. 또한 트레드밀 운동은 파킨슨질환의 병리학적 특징인 α-syn 단백질의 발현을 감소시키고 산화적 스트레스를 완화시키는 항산화 효소들의 활성을 증가시키는 것으로 보고하였다[22]. 따라서 운동에 의한 α-syn 단백질과 산화적 스트레스의 감소는 파킨슨질환에 긍정적인 영향을 미칠 것으로 생각될 뿐만 아니라 미토콘드리아 역동성의 장애를 일부 개선시킬 수 있을 가능성을 제시할 수 있지만 아직까지 파킨슨질환 유도 모델 동물을 대상으로 운동에 의한 α-syn 단백질과 미토콘드리아 역동성의 관계를 살펴본 연구는 매우 부족한 실정이다. 흥미롭게도 미토콘드리아 기능이상이 파킨슨질환의 병리학적 특징으로 보고되면서, 운동에 의한 α-syn 단백질의 감소가 미토콘드리아 기능개선에 미치는 영향을 확인하는 것은 운동이 파킨슨질환에 긍정적인 영향을 미친다는 메커니즘을 구체적으로 제시할 수 있을 것으로 생각된다.
따라서 본 연구의 목적은 1-methyl-1,2,3,6-tetrahydropyridine (MPTP) 약물을 투여한 파킨슨질환 모델 쥐를 대상으로 8주간의 트레드밀 운동을 통해 흑질의 α-syn 단백질 발현과 미토콘드리아 역동성에 미치는 영향을 확인하는 데 있다.
연구 방법
1. 연구 대상
본 연구에서 사용된 실험동물은 C57BL/6 마우스를(n =24, 8주령) 분양 받아 K 대학교 동물사육실[온도 20±2°C, 습도 50%, 주간(08:00-20:00)에 점등, 야간(20:00-08:00)에 소등]에서 사육하였다. 집단은 생리식염수를 투여하는 Sham집단(n = 8, Sham), MPTP 투여집단(n = 8, MPTP) 및 MPTP 투여 후 트레드밀 운동집단(n = 8, MPTP-E)으로 총 3 집단으로 구분하였으며, 실험기간 동안 식이량과 수분섭취는 제한 없이 공급하였다.
2. 연구절차
1) 파킨슨질환 마우스 모델 확립
본 실험에서 만성 파킨슨질환 마우스 모델을 확립하기 위해 복강 내에 25 mg/kg의 1-methyl-1,2,3,6-tetrahydropyridine (MPTP, Sigma-Aldrich)와 흡수를 돕기 위한 항 보강제인 Probenecid (Sigma-Aldrich, 250 mg/kg)을 복강(intraperitoneal)에 총 10회(2회/주, 5주간) 투여하였다. 투여 방법과 절차는 Lau et al. [23]가 제시한 방법을 적용하여 실시하였다.
2) 운동기능 검사(Rota-rod test)
본 연구는 동물모델의 운동협응 기능과 균형감각을 평가하기 위해 Rota-rod test (five onin acclerating retarod; JD-A-07, Jeung do, Korea)를 이용하였다. 본 검사 전 실험동물을 120초 동안 10 rpm의 속도로 회전 원통에서 사전적응훈련을 실시하였으며, 본 검사에서는 회전속도를 5-40 rpm까지 점진적으로 증가시켜 진행하였다. 300초의 최대 수행제한 시간 내에서 실험동물이 떨어지는 시간을 2회 측정하여 평균값을 기록하였다.
3) 트레드밀 운동 프로그램
실험동물은 rodent 트레드밀(8Lanes, Daemyung Scientific Co, Ltd, Korea)을 이용하여 1주간 사전 적응훈련(10 min/day, 10 m/min, 5 days/week)을 실시하였다. 이후 처음 2주 동안(10 m/min, 30 min/day, 5 days/week) 운동을 실시하고 점진적으로 운동 강도를 증가시켜 마지막 8주 동안에는 12 m/min 속도로 60분간 실시하였다. 운동 프로그램은 이들의 운동수행능력을 Lau et al. [23]가 제시한 운동 프로그램을 수정하여 실시하였다.
4) 뇌 적출
8주간 트레드밀 운동과 운동기능검사를 실시한 후 pentobarbital sodium (50 mg/kg)을 복강 내 주입시켜 마취시킨 후, 중뇌 흑질을 적출하여 미토콘드리아를 분리하고 남은 조직은 액화질소에 동결시켜 분석 시까지 -80°C의 초저온 냉동기(Bio-Freezer, Forma Science, USA)에 냉동 보관하였다.
5) 미토콘드리아 분리
미토콘드리아 분리는 Mitochondria Extraction Kit (IMGENEX Corporation, San Diego, CA, USA)을 이용하였다. 분석은 뇌 조직 100 mg 당 1 mL의 homogenizing buffer를 넣어 균질화시킨 후, 4°C에서 10분 간 3,000 rpm으로 원심 분리하고 분리된 상층액을 다시 4°C에서 30분 간 12,000 rpm으로 원심 분리하였다. 원심분리한 상층액(cytosolic fraction)은 세포질로 분리하고 남은 pellet을 1 mL의 suspension buffer를 넣어 잘 섞어 준 다음 다시 4°C에서 10분간 12,000 rpm으로 원심 분리하였다. 이후 상층액을 제거 후 1회 더 suspension buffer 1 mL을 넣고 잘 섞어 준 후 다시 4℃에서 10분간 12,000 rpm으로 원심분리해서 상층액을 제거한 후 남은 pellet은 1 mL의 Complete Mitochondrial lysis buffer를 넣어 4°C에서 30분간 녹인 후 분리된 mitochondrial extract를 4°C에서 5분간 12,000 rpm으로 원심 분리하여 획득한 상층액(mitochondria fraction)을 분리하고 총 단백질량은 BSA (Bovine Serum Albumin, 570 nm)를 이용하여 정량하였다.
6) Western blot
집단 별로 분리된 뇌 조직의 미토콘드리아와 세포질의 총 단백질량을 정량하고 준비된 단백질은 20-30 µg으로 7.5-12% SDS-Polyacrylamide gel에서 전기영동 후 membrane으로 전이시키고, 5% BSA가 첨가된 1×TBS-T 용액으로 90시간 동안 실온에서 blocking 시킨 후, 각각의 1차 항체와 4°C에서(9-12시간) 반응시켰다. 이후 TBS-T 완충액으로 8분간 5회 세척 후 2차 항체와 실온에서 90분 동안 반응시킨 후에 다시 TBS-T 완충액으로 8분간 5회 세척 후, WBLR solution (Western Blotting Luminol Reagent SC-2048, Santacruz Biotechnology, USA)에 membrane을 넣고 1분간 발색하고 얻어진 membrane을 이미지 분석 시스템(Molecular Imager ChemiDoc XRS System, Bio-Rad, USA)을 이용하여 스캔한 후 Quantity One 1-D Analysis Software (Bio-Rad, USA)를 이용하여 단백질량을 분석하였다.
7) Immunofluorescence (IF)
각 집단에서 선택된 뇌 조직 샘플(40 µm)을 자유부유법(Free-floating method)을 이용하여 10 mM PBS로 각 5분씩 3번 세척하고 0.01 M Sodium Citrate buffer에 담아 90°C에서 60분 동안 반응시켜 항원복구 과정(Antigen retrieval)을 진행한 후 PBS로 5분씩 4번 세척하였다. 항원의 비 특이적 결합을 막기 위해 10% Normal donkey serum을 이용하여 실온에서 60분 동안 반응시킨 후 1차 항체인anti-α-synuclein (BD bioscience, bd610787, USA) 단백질을 24시간 동안 4°C에서 반응시켰다. 이후, 조직샘플을 5분 4번씩 세척한 후 실온에서 2시간 동안 2차 항체(cy3 conjugated donkey anti-mouse; 1:200 dilution, Jackson Immunochemicals, West Grove, PA, USA)에 반응시켰다. 4번의 세척과정을 거친 후 각 조직은 슬라이드에 옮겨 마운팅 용액과 함께 봉입하고, immunofluorescence microscope (Leica Microsystems, TCS SP8, Germany) 이용하여 실험결과물을 분석하였다.
연구 결과
1. 트레드밀 운동에 의한 Rota-Rod test 변화
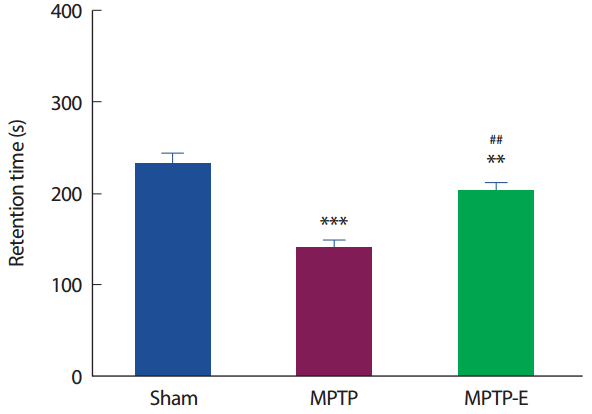
8주간 트레드밀 운동 후 운동기능 능력을 평가하기 위해 Rota-Rod 검사를 실시하였다(Fig. 1). 운동유지 시간은 집단 간 통계적으로 유의한 차이가 나타나 사후검증을 실시한 결과[F(2,21) =24.31, p =.001], MPTP 집단은 Sham 집단과 비교하여 운동유지 시간이 통계적으로 유의하게 감소된 것으로 나타났다(p<.001). 한편, 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 운동유지 시간이 통계적으로 유의하게 증가된 것으로 나타났고(p< .01), Sham 집단과는 유의하게 감소된 것으로 나타났다(p< .01).
2. 트레드밀 운동에 의한 α-syn 단백질과 신경세포사멸의 변화
8주간 트레드밀 운동 후 α-syn 단백질 발현수준을 확인한 결과(Fig. 2B), 집단 간 통계적으로 유의한 차이가 나타나 사후검증을 실시하였다[F(2,12) =24.22, p=.001]. MPTP 집단은 Sham 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났다(p< .001). 반면 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났고(p< .01), Sham 집단과는 유의하게 증가된 것으로 나타났다(p< .05). 면역형광염색 또한 단백질 수준과 유사하게 MPTP 집단에서 광범위하게 염색된 반면 운동을 실시한 MPTP-E 집단에서 감소된 것으로 나타났다(Fig. 2F). 또한 α-syn 단백질에 의한 신경세포사멸 관련 단백질 수준을 확인한 결과(Fig. 2C-E), 집단 간 통계적으로유의한차이가나타나사후검증을실시하였다[Bcl-2: F(2,12)=223.13, p=.001, Bax: F(2,12) =29.18, p=.001, Bcl-2/Bax ratio: F(2,12) =271.27, p=.001]. 먼저 Bcl-2 단백질은(Fig. 2C), MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났다(p < .001). 반면 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났고(p< .001), Sham 집단과는 통계적으로 유의하게 감소된 것으로 나타났다(p< .001). Bax 단백질은(Fig. 2D), MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났다(p< .001). 반면 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났고(p< .01), Sham 집단과는 통계적으로 유의하게 증가된 것으로 나타났다(p<.05). 마지막으로 Bcl-2/Bax ratio (Fig. 2E)는 MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 감소되었지만(p< .001), 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났고(p< .001), Sham 집단과는 통계적으로 유의하게 감소된 것으로 나타났다(p< .001).
3. 트레드밀 운동에 의한 Mfn1, Mfn2 및 Opa1 단백질 변화
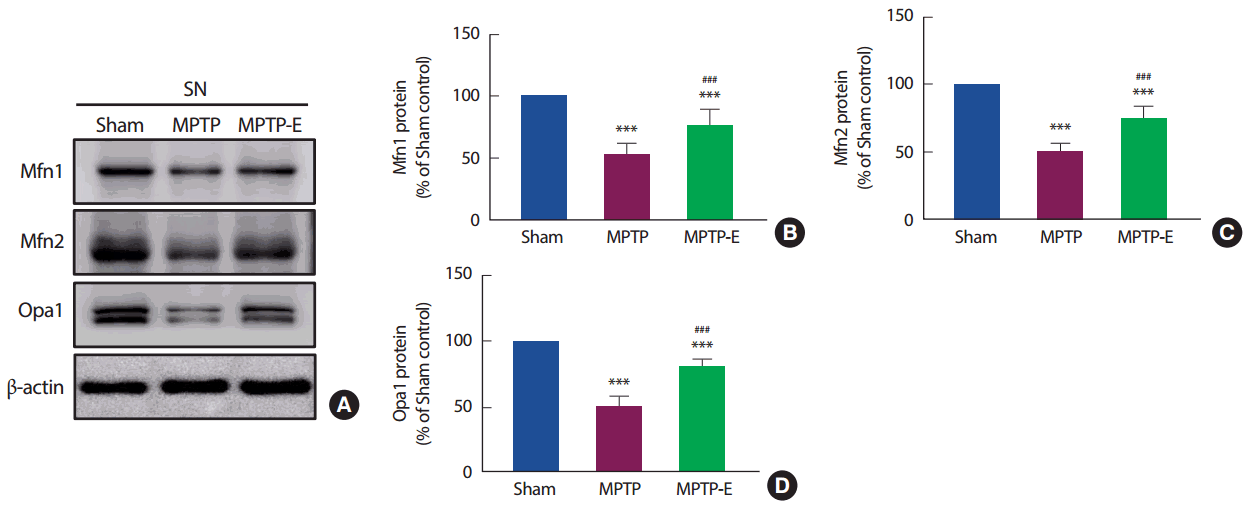
8주간 트레드밀 운동 후 미토콘드리아 합성에 관련된 단백질인 Mfn1, Mfn2 및 Opa1을 확인한 결과(Fig. 3), 집단 간 통계적으로 유의한 차이가 나타나 사후검증을 실시하였다[Mfn1: F(2,12) =796.43, p=.001, Mfn2: F(2,12) =216.54, p=.001, Opa1: F(2,12) =234.99, p=.001]. Mfn1, Mfn2 및 Opa1 단백질 모두(Fig. 3B-D), MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났다(p< .001). 반면, 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났고(p< .001), Sham 집단과는 통계적으로 유의하게 감소된 것으로 나타났다(p< .001).
4. 트레드밀 운동에 의한 Drp1과 Fis1 단백질 변화
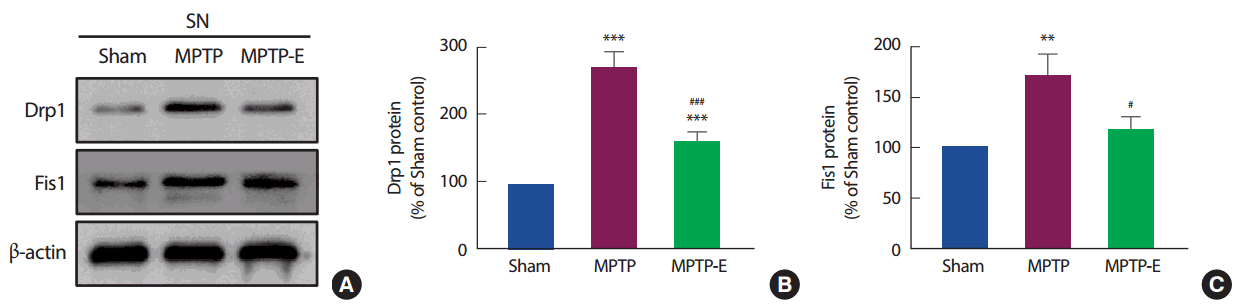
8주간 트레드밀 운동 후 미토콘드리아 분열과 관련된 단백질 수준을 확인한 결과(Fig. 4), 집단 간 통계적으로 유의한 차이가 나타나 사후검증을 실시하였다[Drp1: F(2,12) =130.19, p =.001, Fis1: F(2,12) =7.60, p =.007]. Drp1 단백질은(Fig. 4B), MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났다(p< .001). 반면, 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났고(p< .001), Sham 집단과는 통계적으로 유의하게 증가된 것으로 나타났다(p< .001). 이와 유사하게 Fis1 단백질도(Fig. 4C), MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났지만(p< .01), 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났다(p< .05).
5. 트레드밀 운동에 의한 SIRT-3 단백질 변화
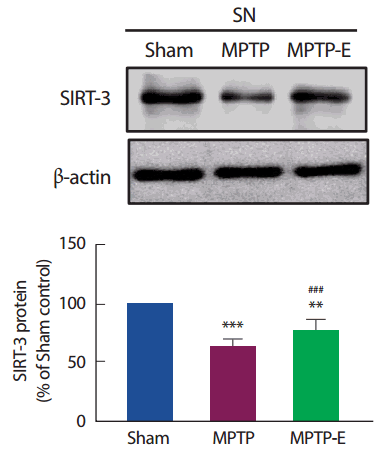
8주간 트레드밀 운동 후 미토콘드리아 SIRT-3 단백질 발현수준을 확인한 결과(Fig. 5), 집단 간 통계적으로 유의한 차이가 나타나 사후검증을 실시하였다[F(2,12) =153.67, p =.001]. SIRT-3 단백질은 MPTP 집단이 Sham 집단과 비교하여 통계적으로 유의하게 감소된 것으로 나타났다(p<.001). 반면, 트레드밀 운동을 실시한 MPTP-E 집단은 MPTP 집단과 비교하여 통계적으로 유의하게 증가된 것으로 나타났지만(p< .001), Sham 집단과는 통계적으로 유의하게 감소된 것으로 나타났다(p< .01).
논 의
1-methyl-1,2,3,6-tetrahydropyridine (MPTP)는 미토콘드리아 전자전달계 복합체 I (complex I)의 기능 억제, 도파민 신경세포 감소 및 α-syn 단백질의 축적을 유도하여 운동장애를 일으키기 때문에 파킨슨질환 유도 모델 쥐의 활용 연구에 사용되는 약물로 알려져 있다[24-26]. 본 연구에서도 운동기능 검사를 실시한 결과 MPTP 집단에서 Rota-rod 유지시간이 유의하게 감소하여 운동기능의 장애가 나타난 것으로 확인되었다(Fig. 1). 이러한 운동기능의 장애는 MPTP 약물 투여로 유도된 것으로 해석되며 이는 동일한 약물을 처치하여 운동기능의 소실을 보고한 선행연구와 일치하는 결과를 나타냈다[27-29]. 반면, 트레드밀 운동을 실시한 MPTP-E 집단에서는 Rota-rod 유지시간이 유의하게 증가되어 운동기능의 장애가 일부 완화된 것으로 나타났으며, 파킨슨질환 모델 쥐를 대상으로 운동 후 운동기능의 회복을 보고한 선행연구와 일치하는 결과를 나타냈다[19,30,31]. 아직까지 운동이 파킨슨질환의 운동기능을 개선시킬 수 있는 메커니즘은 확실하게 밝혀진 바가 없지만 MPTP 약물 처치로 인한 α-syn 단백질의 축적이 신경세포의 사멸과 함께 운동기능의 장애를 증가시킨다고 보고되었다[32,33]. 따라서 α-syn 단백질의 축적을 감소시키는 것이 신경세포사멸을 억제하고 운동장애를 완화시킬 수 있는 방법이라고 생각된다. 예상한 바와 같이 흑질의 α-syn 단백질 발현과 신경세포사멸은 MPTP 집단에서 유의하게 증가된 것으로 나타났고(Fig. 2) 동일한 약물을 처치하여 α-syn 단백질의 축적과 신경세포사멸을 보고한 선행연구와 일치하는 결과를 나타냈다[28,34]. 이는 결과적으로 MPTP 투여로 인한 α-syn 단백질의 축적이 신경세포사멸을 증가시켜 운동장애를 유발한 것으로 판단된다. 반면 8주간의 트레드밀 운동을 실시한 MPTP-E 집단의 경우 α-syn 단백질의 발현과 신경세포사멸이 유의하게 감소되는 것으로 나타났으며, 운동을 통해 α-syn 단백질의 감소를 보고한 Tuon et al. [22]의 선행연구와도 일치하는 결과는 나타냈다. 이를 종합해보면 트레드밀 운동이 α-syn 단백질을 감소시키고 신경세포사멸을 억제시켜 일부 운동기능을 회복시킨 것으로 생각된다. 하지만 아직까지 운동에 의한 α-syn 단백질의 감소를 보고한 연구는 매우 부족한 실정이며, 운동을 통한 α-syn 단백질의 감소 기전도 알려진 바가 없기 때문에 추후 심도 있는 후속연구가 필요하다고 생각된다.
파킨슨질환과 미토콘드리아 기능이상은 매우 밀접하게 관련되어 있으며, 미토콘드리아 기능을 개선시키는 것이 파킨슨질환을 완화시킬 수 있는 새로운 방법으로 제시되고 있다[35,36]. 그 중 미토콘드리아 역동성(mitochondrial dynamic)은 미토콘드리아 구조와 기능을 유지하기 위해 융합과 분열을 끊임없이 반복하여 에너지를 제공하는 기전으로 세포의 생존과정에 매우 중요하다. 미토콘드리아는 mitofusins 1과 2(Mfn1, Mfn2) 및 optic atrophy 1 (Opa1) 단백질을 증가시켜 융합을 유도하고 fission 1 (Fis 1)과 dynamin related protein 1 (Drp 1) 단백질의 발현을 증가시켜 분열을 유도한다[37,38]. 하지만 최근, α-syn 단백질의 축적은 미토콘드리아 융합을 억제하고 분열을 증가시켜 미토콘드리아 구조와 기능을 저하시키는 것으로 보고되었다[12,13]. 본 연구에서도 MPTP 그룹에서 미토콘드리아 융합과 관련된 Mfn1, Mfn2 및 Opa1 단백질이 유의하게 감소된 것으로 나타났고(Fig. 3), 미토콘드리아 분열에 관련된 Drp1과 Fis1 단백질이 유의하게 증가된 것으로 나타났다(Fig. 4). 이는 앞서 제시한 것처럼 MPTP 약물 투여로 인한 α-syn 단백질의 축적이 미토콘드리아 역동성의 불균형을 초래하고 미토콘드리아 분절을 증가시켜 미토콘드리아 기능이상이 나타난 것으로 판단된다. 일반적으로 운동은 미토콘드리아의 생합성(mitochondrial biogenesis), 미토콘드리아 역동성 및 손상된 미토콘드리아를 제거하는 미토파지(mitophagy)를 활성화시켜 미토콘드리아 기능을 개선시키는 것으로 알려져 있다[18]. 하지만, 아직까지 파킨슨 질환을 대상으로 운동을 통해 미토콘드리아 역동성을 확인한 연구는 매우 부족한 실정이다. 이에 본 연구에서살펴본결과트레드밀운동을실시한 MPTP-E 집단에서 Mfn1, Mfn2 및 Opa1 단백질이 증가된 반면, Drp1과 Fis1 단백질은 유의하게 감소되어 미토콘드리아 역동성의 불균형을 일부 개선시킨 것으로 나타났다. 따라서 본 연구에서 나타난 미토콘드리아 역동성의 개선은 일부 운동의 효과라고 생각되며, 이는 파킨슨질환 모델 동물 쥐를 대상으로 운동에 의한 미토콘드리아 역동성의 활성을 확인한 의미있는 연구라고 생각된다.
Sirtuin-3 (SIRT-3) 미토콘드리아에 존재하는 단백질로써 NAD+ 의존성 디아세틸레이션(NAD+ dependent deacetylation)을 통해 지방산화(fat oxidation), 미토콘드리아 생합성(mitochondrial biogenesis) 및 ATP 합성을 증가시킬 뿐만 아니라 산화적 스트레스를 증가시키는 ROS의 생성을 감소시킨다[39,40]. 또한, SIRT-3는 미토콘드리아 융합 단백질인 Opa1의 활성을 증가시키는 반면, 분열 단백질인 Fis1와 Drp1을 감소시키는 것으로 보고되었다[41,42]. 이에 본 연구에서 SIRT-3의 발현을 확인한 결과(Fig. 5), 트레드밀 운동을 실시한 MPTP-E 집단에서 유의하게 증가된 것으로 나타나, 운동을 통해 SIRT-3의 발현이 증가됨을 보고한 여러 선행연구와 일치하는 결과를 나타냈다[43,44]. 이러한 SIRT-3의 증가는 결국 운동 후 미토콘드리아 역동성의 개선이 나타났던 근거를 일부 뒷받침해 줄 수 있는 것으로 생각된다.
따라서 운동은 파킨슨질환의 병리학적 특징인 α-syn 단백질의 축적을 감소시키고 신경세포사멸을 완화시켰다는 것을 확인하였고, 이에 운동기능의 장애가 일부 개선되었다는 것을 보여주었다. 또한, 트레드밀 운동은 SIRT-3 단백질을 증가시키고 미토콘드리아 역동성의 불균형을 완화시켜 파킨슨질환에 나타나는 미토콘드리아 기능이상을 일부 개선시킬 수 있는 효과적인 방법이라고 생각된다.
결 론
본 연구에서는 MPTP 약물을 투여한 파킨슨질환 동물 모델을 대상으로 8주간의 트레드밀 운동이 운동기능, α-syn, 신경세포사멸, 미토콘드리아 역동성 및 SIRT-3 단백질에 미치는 영향을 확인하고 다음과 같은 결론을 도출하였다. 8주간의 트레드밀 운동은 α-syn 단백질의 감소시키고 신경세포사멸을 억제시켜 운동기능 소실을 일부 회복시킨 것으로 나타났다. 또한, 트레드밀 운동은 SIRT-3 단백질을 증가시켜 미토콘드리아 역동성의 불균형을 일부 완화시켰다. 따라서 운동은 파킨슨 질환에 나타나는 미토콘드리아 기능이상을 일부 개선시킬 수 있는 효과적인 방법이라고 생각된다.